


 13720223167
13720223167




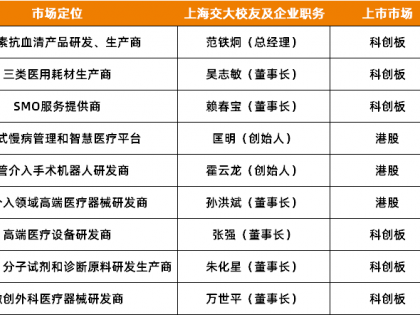
近日,南洋理工大学梁照珣课题组在JACS 上发表了题为“源于非经典烯二炔生物合成途径的Sungeidines”的研究成果,为烯二炔家族新添了一重要成员分子。作者通过基因组导向方法从小单孢菌中发现了一类全新骨架分子Sungeidines,其生物合成基因簇(BGC)与已知的dynemicin类烯二炔存在同源进化关系但又表现出显著的差异。同位素标记实验结合基因信息分析表明,该类分子是由八酮链(octaketide)前体经不同修饰(氧化、环化、断键等)后形成的两个C15骨架组合而成;同时发现并表征了BGC中一个新颖的磺酰基转移酶(SgdX2)介导的脱水反应。作者认为Sungeidines与经典烯二炔之间的结构差异是由于进化过程中缺失了部分的关键修饰酶(如环氧化酶)所致。该研究揭示了烯二炔生物合成途径的可进化性和可塑性,并为进一步探究Sungeidines生物合成中有趣的酶学现象奠定了基础。
作者从新加坡红树林的沉积物中分离得到了一株小单孢菌Micromonospora sp. MD118,基因组测序结果显示其含有一个与dynemicin型烯二炔类似的PKS基因簇,都包括一个聚酮合酶(sgdE8),一个硫酯酶基因(sgdE7)和三个未知基因(sgdT3,U14和U15);而与经典的dynemicin烯二炔相比其不同之处在于,sgd BGC中缺少了一些相对保持的基因(如dynA1, dynA2和dynE13),但同时具有5个特有的基因(sgdX1-X5,图1)。作者采用基于CRISPR/Cas9的基因敲除方法确认了sgd BGC所对应的代谢产物,并经转录调控修饰获得了高产菌株MD118AE,从中成功分离鉴定了主产物成分sungeidines A (1)和B (2), 其结构由一个高度稠合的四环骨架与一个变异的蒽醌骨架骈合而成,与dynemicin相比,最显著区别为sungeidines骨架中多出了两个额外的碳原子(图1)。与此同时,还分离到了一个碘代的蒽并硫酯类化合物9,其结构与sungeidines左单体片段的骨架一致,但遗憾的是,饲喂发酵实验证明9并非为sungeidines生物合成途径中的中间产物。
图1. 基因挖掘下Sungeidines的发现
为探究导致sungeidines和dynemicins结构差异的原因,作者对特有基因sgdX1-X3进行了逐一敲除,结果发现ΔsgdX2突变菌株不再代谢1和2,而其余产物的量显著增加,后经分离鉴定为sungeidines C-H (3-8),其结构中均含有邻二醇结构。为进一步验证SgdX2的生物功能,作者将其在大肠杆菌中进行过表达后纯化得到SgdX2。经体外生化实验证实SgdX2可催化6形成不稳定的磺酸酯中间体,而后可经自发脱水芳化生成化合物1(图2)。
图2. Sungeidines C-H (3-8) 的结构及SgdX2的功能表征
基于上述分析,作者认为sungeidines是由两个聚酮来源的C15骨架组装而成,而dynemicins则由两个C14碳骨架衍生而来。为验证该猜想尤其是额外的两个碳原子的来源,作者设计了同位素示踪实验,并经NMR分析表明sungeidine中所有碳原子均来源于乙酸,C15骨架是由八聚酮类中间体(octaketide)断掉末端碳原子而成(图3)。因此,sungeidines和dynemicins可能经历共同的octaketide前体,而后经不同的后期修饰形成了不同的C14和C15骨架。
图3. Sungeidine的同位素标记实验
此外,作者尝试各种培养条件均未检测到sungeidines的活性烯二炔中间体,推测可能是由于dynA1、dynA2和dynE13(环氧合酶)同源基因的缺失,导致烯二炔中间体缺乏可稳定其构象的必要基团,而易发生环化形成其他结构,据悉环氧化物或环外烯烃基团的存在对于防止十元烯二炔自发的环芳构化反应至关重要。因此,作者认为sungeidines应同样源于dynemicins类烯二炔的生物合成途径,但是在适应性进化过程中缺失了相关的后期修饰基因如环氧合酶而导致主骨架发生变化,表明后修饰基因的干预或定向改造在获取结构丰富多样的代谢产物上具有重要潜力。综上所述,作者提出了sungeidines生源后期的合成通路(图4)并为研究sungeidines中C-I、C-S、C-N和炔键的形成及环化机理打下基础。
图4. 作者提出的生物合成途径




 加载中...
加载中...