


 13720223167
13720223167



单位:教育部合成生物学前沿科学中心,系统生物工程教育部重点实验室,天津大学化工学院
引用:丁明珠, 李炳志, 王颖, 谢泽雄, 刘夺, 元英进. 合成生物学重要研究方向进展[J]. 合成生物学, 2020, 1(1): 7-28
合成生物学作为一个新兴的交叉学科领域,随着DNA合成技术的进步和合成生物学理念的深入,多个研究方向取得了长足发展。本文主要对基因回路、基因组设计合成、细胞工厂和人工多细胞体系的进展进行了综述。可设计构建的人工基因线路的复杂度逐步提升,人工控制更加精细;组装技术取得快速进展的同时,人工基因组的设计深度也在不断拓展,设计合成的人工基因组由支原体拓展向大肠杆菌,甚至真核生物酿酒酵母,推动了生物进化演化的研究;细胞工厂的设计构建在逐步挑战代谢途径更长、复杂程度更高的化合物的合成,模块化和正交化策略对复杂细胞工厂构建的支撑作用日益明显,鲁棒性和适配性成为细胞工厂构建需要考虑的重要问题;人工多细胞体系的设计构建已经从设计构建两菌体系向多菌体系扩展,通过多种原则进行设计,实现更加复杂的预期功能。本文也对合成生物学与其他学科交叉融合产生的一些新研究方向进行了简介。
合成生物学是指在工程学思想指导下,按照特定目标理性设计、改造乃至从头重新合成生物体系,即生物学的工程化。合成生物学的出现是多学科交叉发展的必然。从DNA双螺旋结构的发现与遗传中心法则的阐明开始,到大规模测序技术推动下越来越多基因组遗传信息的解读,人们对生物功能与基因和基因组关系的理解逐渐深刻。工程学思想的引入加速了生物学、生物信息学、计算机科学、化学、材料学等多学科的交叉融合,推动了生物学的工程化从模块化、定量化、标准化、通用性等角度系统展开,形成了合成生物学的研究领域。
DNA的人工合成是合成生物学研究的底层推动技术。DNA化学合成法是当前主流的商业化合成方法,在合成长度和合成通量上不断取得突破,发展出柱式合成和芯片合成工艺,使寡核苷酸链合成通量可以达到百万条级,合成成本降低约3个数量级。近年来,DNA酶促合成法日益受到关注,利用末端脱氧核苷酸转移酶(TdT)在温和的条件进行寡核苷酸链的合成,有望推动DNA合成技术的再次升级。随着DNA合成技术的快速发展,合成生物学在多个研究方向不断取得突破。本文将对基因回路、基因组设计合成、细胞工厂和人工多细胞体系的重要研究进展进行综述,并对合成生物学与其他学科交叉融合产生的一些新研究方向进行简介。
1 基因回路
基因回路是由调控元件和被调控的基因构成的特定逻辑关系,以实现所设计的预期功能。“双稳态开关”(Toggle switch)基因回路及“自激振荡网络”(Oscillatory network)基因回路的研究开启了这一合成生物学基础研究分支。基因回路往往借鉴电器回路的逻辑控制概念,与底盘细胞基因网络形成一定程度的正交关系,其未来的一个重要发展方向是构建“生物计算机”的“编程与控制”系统。基因回路的要素有:标准化的基因元件库、多基因之间的调控逻辑、调谐与降噪等回路整体的维护,最终实现由简单到复杂的目标功能(图1)。由基因回路组装更大规模的网络时,对模块化封装提出需求。此外,当搭载基因回路的个体细胞以群体形式执行功能时,也需要细胞间能够通讯、互作、分工。

图1 基因回路的组成
1.1 标准化的基因元件库
1.2 基因间的调控逻辑与基因回路调谐降噪
基因回路最常用的调控逻辑参考电器元件中的“开关”与“逻辑门”。基因开关包含多种形式,如能在两个基因表达稳态之间切换的双稳态开关、能被特定化学小分子诱导开启的核糖开关、能由RNA变构进行控制的RNA开关等。对于基因逻辑门,可在单个细胞中建立转录级联控制的分层逻辑门,也可利用重组酶的通用特性建立16种“2输入1输出型”布尔逻辑门。基因开关与逻辑门的开发增强了细胞选择与执行复杂行为的能力。
基因回路需要对多个基因表达进行精细控制,才能实现整体的精细功能。利用降解标签来控制元件的蛋白质降解时间,可以设计构建出频率与振幅均可自由调控的振荡器装置。利用逻辑“非门”检查细胞对时序逻辑的执行,从而达到回路时序控制。通过构建“长寿命基因时钟”,可延长基因回路在细胞中发挥功能的时间,实现回路的时限控制。通过调控元件生物分子的降解速率,可将振荡器振荡周期的标准差从35%降低到14%,达到回路降噪的目的。
1.3 基因回路的模块化封装与细胞间互作
当不同的基因回路组成更加复杂的基因调控网络时,需要对回路进行一定程度的模块化封装。利用正交化-模块化病毒蛋白酶可对靶蛋白进行对接与切割,实现了8种二进制逻辑门和动态模拟信号处理等封装功能。将基因回路包裹在脂质体小滴中,添加基因表达所需工具酶等组分,可控制各回路独立工作与产物交流。为了对基因回路的设计进行加密,可分别建立“加密回路拓扑”与“模糊化回路连接”策略,使得未经授权则难以确定回路结构和功能。
搭载回路的多个细胞组成的细胞群可构成协同、竞争等不同的群体关系。“群体感应”基因回路已从原核细胞拓展到真核细胞中。基因回路控制的亚细胞群可形成“捕食者”关系模型、细胞“荧光条纹分布”模型等。通过建立大肠杆菌菌斑之间的通讯,可构建所有双输入布尔逻辑门。通过混养搭载不同基因回路的酵母细胞亚群,可实现多路选择器和进位加法器的逻辑运算。个体细胞之间也可发展成可控的多细胞结构,例如通过建立动物细胞模块化分泌信号平台,可控制细胞表面钙黏蛋白的表达,从而控制多细胞间的黏附结合与整体结构。
1.4 基因回路走向应用
基因回路发展的高阶目标是实现生命功能的程序化控制。根据目标需求,合理设计并组合使用基因回路,可使细胞编程更加精细化,逐步实现预设合成生物功能的高度可控甚至完全可控。
2 人工基因组设计合成
2010年,Synthia细胞的自我复制标志着人工合成基因组实现了对生命活动的调控,突破了化学物质和活性基因组的界限。人工基因组合成是关于理性设计和重新合成生命的研究,即在工程学思想的指导下,借助计算机模拟,模块化设计具有特定功能的人工基因组,利用DNA从头合成和模块化组装技术,将人工设计基因组构建出来,并使其实现预期功能。
近年来,人工基因组合成取得了一系列重大突破。最小化基因组的理性合成颠覆了简化生命体的传统策略,使我们对人工细胞在特定环境下的行为和功能机制的理解更加深入。密码子转换和非天然氨基酸技术的应用实现了正交化生命体的创建,拓展了生命进化方向和生命存在形式的可能性。人工基因组合成的发展内容主要包括DNA片段高效组装和迭代替换、基因组精简与遗传密码扩展、遗传系统可控进化等,最终实现人工细胞性能的定向优化(图2)。
图2 人工基因组的设计、合成与应用
2.1 DNA片段高效组装和迭代替换
人工基因组的长度远远超出了普通DNA化学合成的长度范围,需要通过标准化DNA元件的逐级组装技术来实现。根据组装原理不同,可将DNA元件组装技术分为酶促体外组装(基于DNA聚合酶、核酸内切酶或核酸外切酶)、非酶促体外组装和体内组装(基于DNA同源重组或位点特异性重组)。酶促体外组装和体内组装技术已应用于支原体、大肠杆菌和酿酒酵母的人工基因组合成,推动了组装大片段DNA(kb至Mb级)的发展。
随后,标准化DNA元件在细胞内对天然基因组进行迭代替换,直至获得完整人工基因组。位点特异性重组和同源重组技术分别应用于原核和真核基因组的迭代替换,同时基因编辑技术大幅提高DNA迭代替换的效率。为进一步加速人工基因组合成的速度,不同的DNA元件分别在多个细胞中进行迭代替换,通过细胞融合进行基因组的转移,快速获得具有完整活性的人工基因组。
2.2 基因组精简与密码子扩展
人工基因组合成能力的提升推动了基因组设计深度的扩展,可以进行复杂遗传信息的精简,重塑细胞生命活动。基因组精简的研究在支原体、大肠杆菌和酿酒酵母等多个物种中展开,主要策略包括非必需基因精简、遗传元件的移除或移位、密码子的简化等。在最小支原体基因组的构建中,精简了大量非必需基因和半必需基因,获得的最小支原体基因组长度缩减50%,仅由473个基因组成。在酿酒酵母人工基因组Sc2.0的设计中,删除了转座序列、内含子、亚端粒等遗传元件,对tRNA基因进行重新定位,人工酵母基因组序列精简了6%。基因组合成可以实现全基因组序列的密码子转换,已实现大肠杆菌基因组1.8万个靶标密码子的转换,精简了丝氨酸密码子TCG、TCA和终止密码子TAG,构建了仅有61个遗传密码的人工大肠杆菌。基因编辑以天然的生物基因组为蓝本,可以修饰和改造特定位点的DNA序列,已实现对大肠杆菌基因组314个TAG终止密码子的转换。随着基因编辑效率的提高、脱靶效应的降低和新方法的不断更新,尤其是碱基编辑器技术的发展,可进行密码子转换的基因组DNA序列区域不断扩大,基因编辑将推动基因组设计与合成的快速发展。
2.3 遗传系统进化和演化
人工基因组合成研究显著提升了获得基因组变异的能力,可以为人工细胞进化和演化提供驱动力,为深入探索系统进化和物种演化的分子机制提供了新的平台。人工基因组的进化和演化在原核和真核细胞中开展,主要包括重叠基因的重构、基因组诱导重排等。人工大肠杆菌基因组中79个重叠基因的重构,实现了基因重叠区域的独立转录,探究了基因编码元件交替分布排列的演化特征。人工酵母基因组的诱导重排研究,揭示了特殊环境下的基因组变异规律和适应性演化机制;并利用该技术开展高产植物源化合物的人工酵母底盘细胞构建和功能基因组研究,提升胡萝卜素产量38.8倍,发现决定重要性状的关键基因,阐明基因表达调控、代谢调节等网络的组成和变化。
人工基因组合成的长远目标是理性设计基因组序列,工程化构建生物遗传系统,定制细胞功能。这需要深度融合生物信息、大规模数据、数理科学、化学、计算机科学等知识,跨尺度地研究基因组与细胞不同组分的互作机制,揭示细胞内整体生命活动规律。
3 细胞工厂的设计构建
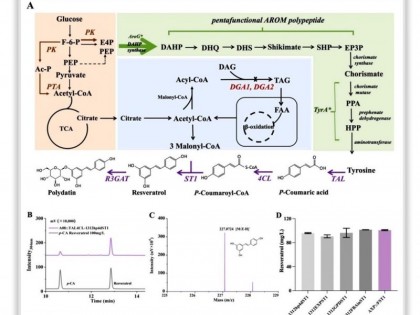
构建细胞工厂从头合成生物基材料单体1,3-丙二醇和重要医药中间体青蒿酸,开创了传统石化产品和天然产物全新的生产模式,可有效解决石油化工炼制和植物提取对自然资源的依赖和对环境的危害。自此,细胞工厂的设计构建成为合成生物学的一个重要研究方向。伴随着合成生物技术的进步,人们在逐步挑战代谢途径更长、复杂程度更高的化合物合成[图3(a)]。尤其是近期阿片类药物和大麻素的从头合成,展示了构建细胞工厂的巨大潜力。细胞工厂的设计构建,是通过对复杂生命体的工程化重构,实现目标产品的可控和高效合成。其实现过程是在物质代谢和能量代谢水平上,反复进行模块与模块、模块与底盘之间的适配。除适配性外,细胞工厂的发展趋势还承袭了模块化、正交性、鲁棒性等工程化特性。
3.1 模块化
代谢途径的模块化是细胞工厂构建的基础。模块中的催化元件或来源于对沉默基因簇和深度测序数据的挖掘,或来源于现有酶分子的定向进化甚至蛋白质的从头设计,以实现自然界中不能催化或难催化的反应,合成非天然的分子,构建未见报道的合成路径。在重构复杂长途径时,采用多元模块工程,按代谢节点和元件功能聚类等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度。通过建立并优化模块功能、调整模块亚细胞器定位以及调节模块之间的表达强度,实现模块与模块之间的组合设计和迭代适配。在上述模块化工程原则的指导下,成功实现阿片类药物、维生素B 12 、托品烷生物碱等长途径的构建。
以途径模块作为基本调控单元,通过调整模块间表达强度,并结合底物通道作用和区室化作用,可以平衡途径上下游或竞争性模块间的代谢流通量,避免代谢中间体和副产物的积累。对模块的表达强度调节,除进行转录水平的调控外,还可进行翻译及翻译后水平的调控。例如在酿酒酵母中通过C-PEST介导的内质网相关降解和N-degron介导的泛素化降解调控蛋白的半衰期,以调整代谢流分配。
图3 细胞工厂的设计构建
3.2 正交化
细胞工厂的正交化设计是指对生产菌株的工程化改造,不干扰底盘原有的代谢和调控网络,以减少人工设计的复杂性。最初正交化设计主要针对导入的异源人工元器件,例如转录因子、核糖体开关和Cas9蛋白等,以实现底盘的基因编辑和表达调控。目前正交化的概念随细胞工厂自下而上的构建过程顺应延伸,逐步扩展到对代谢途径和真核底盘细胞器的设计中。
3.2.1 代谢途径与底盘的相对正交化
在物质代谢水平,代谢途径的相对正交化是指底物同化途径或产物合成途径与底盘内源代谢网络最多只以唯一的结点相连,以形成独立的模块,便于后续调控其代谢流通量。此种途径的正交化过程可视作对底盘生长和产物合成的解耦。在碳源利用方面,常设计构建非天然途径,或利用葡萄糖、油脂等天然碳源[图3(c),正交化途径1],或转化半纤维素水解产物(木糖、阿拉伯糖、半乳糖)等替代碳源[图3(c),正交化途径2],合成关键前体化合物。例如,合成1,4-丁二醇和琥珀酸的正交化途径。在前体供给方面,常基于非天然前体构建目标产品的正交化人工合成途径[图3(c),正交化途径3]。例如,以NPP(neryl diphosphate)替代其异构体GPP作为单萜合成的前体,通过NPP合酶的引入和单萜合酶的定向进化,可构建正交化的橙花醇、桧烯、柠檬烯等单萜类产品的合成途径。
物质转化和能量转移高度耦合。而天然辅酶同时参与众多代谢反应和生物过程,其具有选择性差、生物学效应可预见性低等问题,降低了所涉及途径的正交性。通过设计构建烟酰胺胞嘧啶二核苷酸等非天然辅酶,建立与底盘正交的氧化还原体系,可以从能量代谢的角度增强目标途径与内源代谢网络的正交化程度。
3.2.2 途径定位细胞器的正交化发展趋势
与原核底盘相比,酿酒酵母等真核底盘具备多种可用于途径模块定位的细胞器,产生区室化效应,提高目标产物的合成水平。通过调控细胞器的形态、数量、大小等表型,可进一步提高定位途径的通量。如果靶向的细胞器具备正交性,可降低此种调控对其他细胞器及底盘内源代谢的干扰。根据参与中心代谢的程度,天然细胞器中过氧化物酶体的正交性最高,完全删除过氧化物酶体蛋白不会影响菌体以葡萄糖为底物的生长。而建立人工过氧化物酶体蛋白转运系统,在目标蛋白靶向定位的同时,抑制天然过氧化物酶体蛋白的定位表达,开启了定制化细胞器的研究[图3(d)]。此外,借助低密度膜结合蛋白精确控制囊泡膜的自发弯曲,有望实现不依赖于底盘内源基因调控膜的分裂,而控制细胞器的表型。
原核生物中天然存在由选择性通透的蛋白质衣壳代替磷脂层分子包裹形成的微室。在真核底盘中重建原核微室或构建类微室结构的人工细胞器,进行异源催化反应,也是细胞器正交化的策略手段[图3(d)]。在重构原核微室时,采用SpyTag/SpyCatcher共价结合或CC-Di-A/CC-Di-B卷曲螺旋(coiled-coil)等蛋白质自主装结构,替代天然微室的包装序列,可解决催化蛋白易发生团聚而丧失活性的问题。但上述方法尚未在真核底盘中进行尝试。而参考天然微室结构,利用自组装蛋白(例如encapsulins、受蓝光调控的Cry2和PixD/PixE)的可控表达和组装,已成功在真核生物中构建了包装有催化蛋白的人工细胞器,实现norcoclaurine、脱氧紫色杆菌素等化合物的高效合成。
3.3 鲁棒性
细胞工厂的鲁棒性是指人工合成体系在受到环境变化和遗传变异等不确定因素干扰时保持其表型稳定的一种特性。这种外界的扰动一方面来源于异源合成途径的引入而产生的代谢负担,以及中间代谢体或终产物积累而带来的胁迫压力和细胞毒性;另一方面来源于实际工业生产过程中的环境压力。下述两种策略分别针对上述两种外界扰动,展示如何增强人工合成体系的鲁棒性。
3.3.1 动态调控合成途径
3.3.2 驯化具有高耐受性的底盘菌株
为应对复杂、非理想化的工业发酵过程,需要底盘菌株拥有抗高温、耐盐、耐酸、耐有机溶剂等性状,以具备对某一类特定环境压力较高的鲁棒性。由于产生耐受性性状具有多基因性,且分子机制并未完全清晰阐释,现阶段很难通过工程化手段完整复制到大肠杆菌和酿酒酵母等常用模式底盘生物中。因此,多采用实验室适应性进化等手段改善模式菌株的耐受性,并挖掘应对相关环境压力的基因组扰动靶点。近年来则倾向于选择天然对极端环境具有耐受性的非模式生物,逐步驯化为可用于细胞工厂构建的底盘菌株。例如耐热耐酸的马克斯克鲁维酵母和溶剂耐受的丙酮丁醇梭菌。以嗜盐假单胞菌为底盘菌株,利用海水为介质高效生产聚羟基脂肪酸酯(PHA)等生物单体材料,是非模式底盘菌株工业应用的典型案例。
综上所述,在细胞工厂的设计构建过程中,模块化和正交化策略的支撑作用日益明显,鲁棒性和适配性成为构建人工合成体系时需要考虑的重要问题。从长远看,细胞工厂将与数学、计算机学、物理学等学科深入交叉融合,实现从原料到菌种再到过程的全链条设计和优化的智能化和自动化。
4 人工多细胞体系的构建与应用
有目标的设计、改造乃至重新合成微生物多细胞体系,在维持菌群结构与功能稳定性、鲁棒性和可控性的同时,可实现复杂的生物功能,提高代谢效率。人工多细胞体系在完成多项复杂任务方面优势明显,减轻单个底盘细胞的代谢负担,同时降低中间代谢物的过度积累和毒害,避免功能间的交叉影响,对环境波动具有更强的适应性和鲁棒性。因此,人工多细胞体系已成为合成生物学研究的重要方向。
4.1 人工多细胞体系的设计构建
人工多细胞体系的设计构建首要解决的问题是,依据什么样的原理和方法设计、构建特定的多细胞体系;其次是如何使各细胞间的功能协调运行,从而实现底物原料到目标产物的高效转化[图4(a)]。
图4 人工多细胞体系的构建与调控
4.1.1 微生物间的互作关系
依据微生物间的互作关系设计构建人工多细胞体系,进一步借助代谢物互补、克服从底物到产物的能量壁垒、解除环境中的抑制作用、调控电子转移平衡等方式,使体系的物质流、能量流和信息流达到适配,可实现人工多细胞体系的高效稳定。细胞间互作关系的稳定存在,普遍依赖于细胞间通讯,细胞间通讯系统是构建人工多细胞体系的关键。研究者通过建立一种预测多细胞体系中相互作用关系的方法,可有效预测800多种菌群中互作关系。哈佛大学研究人员关注由细胞群体感应信号或代谢互补联系起来的混菌共生关系。利用群体感应系统,可构建能够双向通讯的多细胞体系。
自然条件下的多细胞体系,常形成互利、偏利、偏害、竞争等多种相互作用关系。自然形成或人工复配的多细胞体系中一般存在成员分工不明确、代谢水平不平衡、竞争与协作共存等问题,相互作用关系错综复杂。采用模块化构建的方法,可针对性地设计和重构各个功能菌种,构建菌株间的协作关系,使其有利于多细胞体系的高效性和稳定性。
在互利共生体系中,菌株之间共享或者交换营养物质,彼此之间可共同完成某一种化合物的彻底代谢,并分别从中获益。通过构建了营养缺陷型菌株,可以构建互利共生的多细胞系统,使菌株促进彼此的生长。研究者利用双向通讯在两株大肠杆菌中构建了共生系统。在偏利或偏害关系的体系中,对一方菌株有益/有害,另一方则不受影响。菌株间对环境资源(如营养物质、空间等)的争夺则形成了菌株间的竞争关系。在构建人工多细胞体系时,也应充分考虑菌株间竞争、偏利或偏害等关系存在的可能性。部分菌株可能加速对营养物质的吸收,并对这些物质进行与构建目标不相关的无效转化,或者产生对其他菌株有抑制作用的物质,这必然会影响菌群的高效和稳定。通过对混菌体系中菌株关系的重构,可解除竞争抑制,使菌株间偏利共生和竞争关系转变为互利共生。通过人工构建和调控菌株间的互利共生关系,使人工多细胞体系对复杂功能网络进行有效的分工协作,可实现混菌系统的高效、稳定、可控。此外,研究者还设计构建了多种菌群相互作用关系,为构建人工多细胞体系提供新的思路。例如:在两株大肠杆菌中分别构建群体感应信号调控下的细胞自杀基因表达及其抑制途径,可形成捕食者与被捕食者的关系。
依据劳动分工原则构建人工多细胞体系,可将复杂的任务合理分配给不同的菌株,在一定程度上避免单菌中细胞代谢负荷重等问题,在提高效率等方面具有显著优势。依据“劳动分工合作”的原则,构建出大肠杆菌-枯草芽孢杆菌-希瓦氏菌人工三菌产电体系,显著提高了化学能到电能的转化效率;通过将紫杉醇前体(5α-乙酰化-10β-羟化紫杉二烯)的合成途径分工构建到大肠杆菌-酿酒酵母人工两菌体系中,实现了目标产物的高效合成;为实现黄酮类化合物的合成,研究者设计构建大肠杆菌-大肠杆菌双菌体系,显著降低了单菌的代谢负担,使黄烷-3-醇的产量比单菌体系提高970倍。
4.1.2 菌株间的互作方式
人工多细胞体系中菌株间的互作交流主要有两种方式:一种是依赖于微生物细胞与细胞间的直接接触作用方式,主要包括膜囊泡、纳米管和瞬时融合外膜。如通过构建不动杆菌-大肠杆菌互利共生混菌体系研究两菌的代谢交流,发现两菌通过细菌纳米管进行物质交换和信息交流。另一种是菌株可通过间接接触进行物质和能量交换,以维持菌群的生态稳定性。间接接触主要依赖于释放到环境中的化学分子(蛋白质、代谢物等)和电子等。如可利用乙醛的挥发特性作为信号分子,构建不同菌株的共生关系等。
4.1.3 人工两菌体系到人工多菌体系
人工多细胞体系的设计构建已经从两菌体系扩展到多菌体系[图4(b)]。随着菌群中菌株种类的增加,菌株的互作关系愈加复杂,体系的高效性和稳定性会受影响。人工多细胞体系在复杂环境中的鲁棒性研究也是关键问题之一。在构建人工多细胞体系时,更要关注对菌株关系的解析和调控,使多菌分工明确,高效协作。通过控制代谢流分配、减少能量转化过程中的损失、增加功能冗余等策略,使体系内的每一个个体都能够在生产能力、稳定性和鲁棒性方面体现出最优性能。
近年来,人工三菌、四菌体系的构建和应用研究也取得重要进展。通过调控电子载体的合成、碳源的优化分配和利用,构建了高效稳定的人工三菌产电体系,能够利用0.28g葡萄糖稳定产电超过15天。通过构建和调控大肠杆菌三菌体系,减少副产物的形成,实现了木质素单体的高效合成。通过在四株大肠杆菌中表达15个外源基因,将天然产物花青素的合成途径分工构建到大肠杆菌四菌体系,首次实现了以葡萄糖、甘油和木糖为底物合成花青素。
4.2 人工多细胞体系的应用
近年来,人工合成多细胞体系的设计构建取得了重要研究进展,微生物混菌体系在资源能源、环境和人类健康领域等表现出强大的应用前景。
在合成化学品、高附加值产品等方面,研究者通过构建大肠杆菌混菌体系,实现了黏康酸、红景天苷、丁醇、吡喃花青素、黄酮类化合物、苯酚等高效合成。如人工构建的大肠杆菌-大肠杆菌“营养互补”体系可以代谢葡萄糖和木糖生产红景天苷,通过两菌株的分工合成,红景天苷的混菌发酵单位达到6.03g/L,是单菌产量的20倍。
在纤维素等复杂化合物的降解方面,通过调控两菌体系的菌群关系和菌群结构,实现了高效转化木质纤维素生产丁醇和富马酸;通过在酵母表面展示纤维小体形成四菌体系,有效利用纤维素生产乙醇;通过增强“营养-解毒”关系,构建了双梭菌互利共生体系,可将玉米芯转化为丁醇,溶剂终浓度达到22g/L。
人工多细胞体系在环境污染物降解、土壤修复等方面也表现出巨大潜力。人工构建的由两株工程化大肠杆菌构成的混菌体系,实现了降解有机磷杀虫剂。人工构建的大肠杆菌-苍白杆菌混菌体系,可实现对二甲基有机磷的完全矿化。通过构建两株假单胞菌人工混菌体系,可提高对石油硫化物的脱硫作用。人工构建的四菌体系可用于降解碳氢化合物,如常见的原料(橄榄油、石蜡油等)和多芳烃污染物(萘和蒽)等,同时生产表面活性剂。
5 合成生物学与其他学科交叉融合
合成生物学从诞生起始就具有多学科交叉的特点,随着近年合成生物学的发展,与其他学科交叉融合又产生了一些新的方向,如DNA信息存储、DNA折纸、非天然氨基酸、非天然核酸等。
5.1 DNA信息存储
手指、石头、刻痕、绳结在古代被用来记录数据和信息,现在大量的信息主要存储于基于电、光和磁的存储介质中。随着社会发展,不断增长的数据存储需求面临着密度、功耗、体积等诸多挑战。受DNA存储遗传信息启发,人造DNA序列被认为是信息存储的新策略。DNA信息存储是先将数据进行编码转换为DNA序列,借助DNA合成数据存入DNA分子;信息读取是通过高通量测序和信息处理将DNA的分子序列转换为原始存储的数据。DNA存储的核心优势是:①载体特征尺度小,信息存储密度高。例如2017年,研究者利用数字信息传输中的喷泉编码实现了平均千分子DNA拷贝的数据存储与读取,实现了218 PB/g(1PB=10 15 字节)的存储密度。微软等进一步优化信息存储DNA读取中PCR参数,实现了存储密度的进一步提升。②DNA可常温保存,维护成本显著降低,且保存时间久。瑞士苏黎世理工的研究者将DNA封存在特殊的材料中,有望存储千年甚至万年。
DNA存储作为一种新型数据存储方式,也面临很多发展挑战。一是传统的DNA合成技术主要针对生物学研究与应用建立,合成速度较低,成本较高,合成序列的长度与通量等也与数据存储需求存在很大差距。由于目前序列合成的成本远高于测序成本,DNA存储特别适合大量冷数据的长期存储。二是存储在DNA中的数据的读取灵活性仍有待提升,基于目前测序技术的读出方法面临数据恢复可靠性、仪器形态、环境适应性等诸多挑战。2019年微软开发了从写到读的端到端原型系统,距离现代信息存储系统仍有巨大发展空间。三是DNA信息存储与现有电子信息存储之间的编码转化尚不成熟。DNA作为一种物质介质具有各种自身的特点,如有断裂、突变、缺失等风险,近年来各种编码方法在DNA存储的密度、可靠性等方面有所突破。最后,DNA存储的技术链条较长,亟需生物、信息、仪器、机械和工程等领域的整合。
5.2 DNA折纸
DNA作为纳米分子还可以被当作材料的构筑单元。DNA本身的特异性碱基互补配对为特异性组装材料提供了基础,可以构筑出以DNA作为框架的精细纳米结构。DNA折纸就是利用DNA的折叠和自组装形成复杂结构的技术,人工设计一系列DNA片段,借助碱基互补配对原则,可以折叠形成不同的预期几何结构,如三角形、矩形、菱形、五角星及笑脸等精细二维结构。我国学者利用人工设计DNA构建了微型的中国地图。其后,研究者还采用DNA折纸技术制造了齿轮、纳米盒子和花瓶等精细三维结构。由于采用了“自下而上”的理性组装方式,DNA折纸技术在纳米材料、分析科学、医药学、纳米机器人和信息加密等方面均有不同的创新性应用。特殊的DNA折纸结构载体可以用于包裹纳米金造影剂,显著提高成像质量,实现对肿瘤内部结构成像,极大减少造影剂的用量。DNA折纸技术还可以用于分子加密系统,性能可以超越基于硅基计算机的常规加密体系,且同时具有保护信息完整性和访问控制的功能。研究人员利用DNA折纸技术的精细可控性能,创建了迄今为止最快、最持久的DNA纳米马达。
5.3 非天然氨基酸
天然蛋白质由20种天然氨基酸组成,迄今已有超过150种非天然氨基酸可以引入生物体内,其中大多数非天然氨基酸是天然氨基酸的衍生物。非天然氨基酸中根据目的需求可以引入烯基、酮基、醛基、酰氨基等功能基团,调控蛋白质的结构与功能。非天然氨基酸通过对特定蛋白质的修饰,可在生物催化、人工生物防逃逸、蛋白质药物、生物检测等方面有重要的应用。氟色氨酸的引入不仅可以提升转氨酶的活性,还提升了酶的溶剂耐受性和热稳定性。对氨基苯丙氨酸引入到转录因子蛋白LmrR可以将其改变为具有催化活性的腙和肟合成酶。此外,将非天然氨基酸引入大肠杆菌的必需蛋白,可以使大肠杆菌依赖于非天然氨基酸存活,从而达到人工生物体的生物安全防逃逸控制。卤代酪氨酸引入肌红蛋白氧化酶,可以形成光谱探针,为研究酶的催化机制提供了便捷手段。
遗传密码拓展技术是非天然氨基酸引入的重要手段,主要是通过控制终止密码子的读取,进而通过氨酰-tRNA合成酶/tRNA对的改造实现。近年来基因组密码子删减研究可以为遗传密码拓展提供高效的人工底盘细胞,将明显提升引入非天然氨基酸的数量、种类和效率。此外,非天然核酸加入基因组的研究也将为更多非天然氨基酸的进一步高效加入提供新的方向。
5.4 非天然核酸
针对核酸的碱基、磷酸和五碳糖三个主要组成部分,可以分别进行人工修饰或替换,形成非天然核酸(XNA)这样一类核酸分子类似物。XNA能够通过碱基修饰或骨架修饰来拓展遗传信息的存储能力,可以具有部分或全部天然核酸的属性。生物体系引入XNA能够拓展遗传信息的多样性,进而拓展生物性能。非天然碱基研究近年来取得了一些重大突破,如八碱基DNA和具有生命活性的六碱基DNA等。通过对天然碱基(A、T、C、G)的分子结构进行修饰,创造出了两对新的可以互补配对的碱基(S、B、P、Z,其中S和B配对、P和Z配对)。将人工碱基与天然碱基相结合,可以获得含有八种碱基的DNA。研究者在探索非天然碱基多样性的同时,也在尝试使非天然碱基具有生物活性。美国学者在大肠杆菌里成功表达了含有两个非天然碱基编码非天然氨基酸的荧光蛋白,实现了非天然碱基的转录活性,首次实现含六碱基DNA的半人工合成生命。XNA种类的增加不仅提升了遗传信息的密度,还为拓展生物的多样性奠定了基础。
目前XNA主要通过外源添加方式进入生物体,将来开发生物体内合成XNA,将进一步拓展XNA在合成生物学领域的广泛应用。由于XNA在体内的稳定性和特异性有其自身特点,在合成生物学的精准检测和正交控制等方面具有重要的应用潜力。
5.5 生物与材料耦合系统
利用生物酶与人工材料结合,形成具有部分生命特征的耦合系统是当前的一个研究热点。其中,模拟自然界光合作用,将生物酶与半导体光催化剂结合,构建基于酶-光耦联的人工光合系统发展迅速。与自然界光合作用相比,酶-光耦联系统既保留了生物酶的催化能力,又结合了半导体的光吸收能力,通过二者体外灵活组合,打破了自然进化的限制,实现太阳能驱动能源化学品和高附加值化学品的绿色、可持续合成。酸脱氢酶、醛脱氢酶和醇脱氢酶与α-Fe 2 O 3 /BiFeO 3 半导体光电极耦合,实现了太阳能驱动二氧化碳到甲醇还原。光系统(PS Ⅱ)和氢酶与半导体光电极耦合,实现了无偏压下的全水分解。此外,半导体与生命单元的结合还可延伸至细胞,将半导体分别沉积于原核微生物和真核微生物细胞表面,实现了微生物胞内NADPH/NADP + 比例及代谢途径的调控,突破了天然微生物的生产能力上限。为减少纳米材料与细胞膜直接接触对微生物产生的破坏作用,细菌生物被膜可用于半导体大规模锚定,构建生物化学兼容的生物-无机界面。该体系不仅可以发展可循环和可再生的催化反应体系,并且能与胞内的氢化酶耦合实现稳定的光催化产氢。半导体材料与不同酶及不同细胞的兼容组合,打破了自然界进化的限制,极大促进了人工光合领域的发展。
5.6 无细胞蛋白合成
无细胞蛋白合成系统是在试管内重现生命中心法则的全部生化反应,可以重建自然并超越自然的生物分子系统。无细胞蛋白合成系统(cell-free)有助于实现对蛋白质表达体系的高效优化与控制,是合成理化性质复杂的蛋白质的重要方法,如生理毒性蛋白、复杂膜蛋白等。该系统也可以辅助对蛋白质功能的筛选与检测,如将cell-free合成系统与CRISPR-Cas系统相结合来筛选核酸酶。cell-free合成系统能够为活性抗体药物的表达与筛选提供便利,大大缩短活性抗体的研究周期。利用真核生物无细胞系统TNTT7快速体外表达FhSAP2蛋白,有效保持了其主要的构象和抗原性质。通过cell-free系统在天然蛋白质结构中插入非天然氨基酸也是近年来各种基础和应用科学的研究热点。cell-free合成系统可以为抗体偶联药物制备偶联药物前体,消除了内源性氨基酸残基随机生成的偶联物所产生的异质性和不稳定性。在检测方面,cell-free合成系统可以为蛋白质共振配位提供多样性的标记策略,制备用于液相核磁共振波谱分析的蛋白质样品。各种cell-free合成系统的开发使得廉价和快速的重组蛋白表达成为可能,并且各种方法之间能够为表达来自不同生物体的蛋白质进行替代和互补。在多样的cell-free合成系统中,基于大肠杆菌无细胞合成系统得到广泛的研究与应用。基于大肠杆菌RFzero-iy菌株的S30无细胞合成系统实现了高达90%以上的3-碘-L-酪氨酸的插入效率。鉴于系统的灵活性,无细胞蛋白质合成系统已经成为合成生物学的重要工具。
6 结 语
合成生物学各个方向的研究日益活跃,对生物学领域研究的支撑作用日益突出,充分表现出合成生物学的强大创新力。同时,合成生物学将生物学与其他学科进行深度交叉融合,充分发挥定量、设计、工程化等特征,正在发展成为一个基础性学科和工具性学科。将来通过进一步借助自动化、信息化的手段,合成生物学将迎来更快速地发展,将对更多的领域产生重要影响和推动,对解决人类社会发展面临的重大难题提供全新的解决方案。
致谢:在撰写本文过程中,中国科学院深圳先进技术研究院钟超研究员,天津大学齐浩教授、陈为刚副教授和石家福副教授提供了帮助和支持,在此一并致谢。




 加载中...
加载中...