


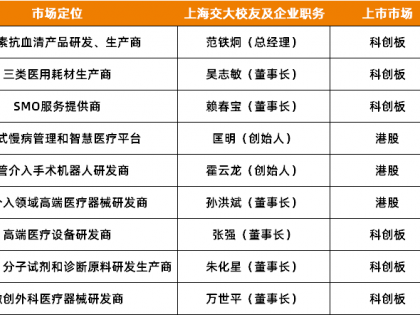
 13720223167
13720223167



IgG是免疫球蛋白中含量最多、研究最深入的一种,Porter早在上世纪60年代就提出IgG的基本结构由4条多肽组成,两条相同的长链称为重链(H链),通过二硫键相互连接构成Y字型骨架;两条相同的短链称为轻链(L链),对称分布在重链两侧。
无论是轻链还是重链都分为稳定区(C区,包括CH和HL,见图)和可变区(V区,包括VH和VL),稳定区的氨基酸数量、种类、排序和蛋白修饰均较为恒定,而可变区则因抗体种类的不同而有所变化,可变区的多样性决定了抗体的多样性与特异性,使得抗体具有结合特定抗原的能力。
随着研究的深入,人们对抗体的了解也越发丰富,相关的研究成果则为后来单克隆抗体的研发奠定了基础。
表1:十九世纪以来抗体领域的重要研究成果:
在抗体应用方面,除了依靠疫苗使人体产生抗体进行治疗的主动免疫以外,人们还通过提取血清中的抗体注射给患者来达到被动免疫的效果。这种通过接种抗原所获得的免疫血清是多种抗体的混合物,它是由多种抗原决定簇刺激多株B细胞增殖分化所产生的,故称为多克隆抗体(多抗)。目前,多克隆抗体主要应用于免疫学检测、被动免疫治疗和紧急预防,但由于多克隆抗体由多种针对不同抗原的抗体组成,因而多抗的特异性差,使用时容易出现交叉免疫反应,故治疗疾病范围有限,也为单抗留下了巨大的发展空间。
长期以来,制备单抗一直是免疫学家追求的梦想,但其中却有一个难以克服的障碍:能够大量产生抗体的浆细胞(plasmacell)作为正常体细胞的一种,具有生长缓慢、无法无限繁殖的缺点,因而无法应用于大规模生产。直到1975年Kohler和Milstein发明了杂交瘤技术才解决了这一难题。
杂交瘤技术的技术流程是这样的:先从被抗原免疫过的小鼠脾脏中提取浆细胞,同时从小鼠骨髓瘤细胞中筛选HGPRT(次黄嘌呤鸟嘌呤磷酸核苷转移酶)缺陷型细胞,将两者在体外进行细胞融合。之后将细胞放入特殊的HAT(次黄嘌呤-氨基蝶呤-胸苷)培养基中进行筛选,未发生融合的浆细胞和骨髓瘤细胞均不能在HAT培养基中生存,从而筛选出了发生融合杂交瘤细胞,该细胞既具有浆细胞的抗体表达能力,又能像肿瘤细胞一样快速生长并无限分裂繁殖,因而可以用于大规模的抗体生产,两位科学家也因此分享了1984年诺贝尔生理学及医学奖。
图1:杂交瘤技术流程图
由于早期单抗生产使用小鼠细胞,产生的鼠源单抗与人源抗体在氨基酸序列、糖基化修饰以及空间结构上有所不同,在进入人体后会被免疫系统认定为外源性蛋白。免疫系统因此产生的鼠源单抗为免疫目标的抗体(即人抗鼠抗体,humananti-mouseantibody,HAMA)不仅会中和鼠源单抗,缩短药物半衰期,影响药效,更会引发免疫反应,轻则出现红疹、风团;重则导致休克甚至威胁生命。因此如何对鼠源单抗进行改造,提高人源化比例,减少HAMA反应便成为了各家单抗生产商的重中之重。
鼠源抗体人源化就是通过基因改造,使单抗和人体内的抗体分子具有尽可能相似的特性,从而逃避人免疫系统的识别,避免HAMA反应。对鼠源抗体进行人源化改造时要遵守两个原则:首先要保持抗体的亲和力和特异性,其次要降低或消除抗体的免疫原性。
嵌合单抗:前文中我们提到抗体识别抗原的能力主要由可变区(V区)决定,因此80年代开始,科学家就希望在保留鼠源V区的同时用人源恒定区代替鼠源的恒定区。这样构建的嵌合抗体既保留了抗体的特异性,又降低了免疫原性。后来成为单抗类第一个明星药物的Rituxan使用的就是这种技术。
人源化单抗:为了进一步减少鼠源成分,90年代后期上市的人源化抗体在保留V区中决定抗体亲和力和特异性的最重要部分——互补决定区(CDR)的前提下用人源的V区替换掉鼠源的V区,就可以使单抗的人源化比例提高到90%-95%,从而使人源化单抗副作用进一步减少。
表2:人源化比例的上升带来不良反应比例的下降
全人源化单抗:随着单抗市场的不断拓展和成熟,产业界对于全人源化单抗的需求也越来越迫切。1993年剑桥抗体技术(CAT)和巴斯夫生物研究公司(后为Abbott收购)开始了第一个全人单抗Humira(抗体名Adalimumab)的研发,有赖于CAT的噬菌体展示技术(Phagedisplaytechnology),Humira于2002年作为治疗类风湿的特效药上市,年销售额短短4年内超过20亿美元,成为单抗领域新的重磅炸弹,也拉开了单抗全人源化的浪潮。
噬菌体是专门感染细菌的病毒,噬菌体展示技术即用多聚酶链式反应(PCR)从生物体内扩增出整套编码人抗体的基因序列,克隆到噬菌体载体上,并通过细菌进行复制,构建噬菌体文库(PhageLibrary)。这些抗体基因会以融合蛋白的形式表达到噬菌体表面,如果将这些带有抗体的噬菌体和含有抗原靶位的细胞混合,就可以方便地利用抗原-抗体特异性结合反应筛选出与抗原结合最为紧密的抗体。通过对此类抗体的进行优化改造即可得到期望中的单抗药物。此项技术不仅可以获得全人单抗,而且不需要细胞融合,不经过免疫动物,实验周期短,过程简单,这是人源抗体制备技术的重大突破,目前国际上主流单抗生产企业均使用该技术筛选单抗药物。
图2:使用噬菌体展示技术筛选全人单抗药物
单抗在筛选完成后下一步便是抗体生产,即单抗的表达。因为单抗分子量较大,每次治疗用药量一般均达到几百mg(小分子化学药用药量一般为几到几十mg),这就对单抗生产商的表达培养能力提出了较高的要求。而判断一家企业生产水平高低的指标主要有表达方法、反应器规模、表达体系和表达量这四要素。
表达方法:早期的单抗生产是将筛选出的杂交瘤细胞植入免疫缺陷型小鼠体内任其生长,抽取小鼠的腹水提取单抗,这种方法称为体内培养法。由于该方法操作复杂,耗时长,后来逐渐为杂交瘤细胞体外培养法取代,而近年来随着基因工程技术的发展,可以直接将编码单抗的基因转入工程菌株/细胞株中,通过工程菌株/细胞株大量表达单抗,降低生产成本。
反应器规模:任何依赖发酵表达的生物药都有着明显的规模效应,即在同等表达量水平下反应器(发酵罐)的容量越大,其单位时间内的药物产出量也就越大,目前国际上的单抗细胞反应器也有持续大型化的趋势,从早期的500L拓展到千L级,大的单体发酵罐甚至达到10000L-25000L,而最大单抗生产企业Genentech的发酵罐总规模已经达到330,000L,占到全球工业用细胞培养能力的24%。
图3:工业化单抗生产流程
表达体系:表达体系指的是使用怎样类型的工程菌株/细胞株来生产单抗,目前常用的人源化抗体表达系统主要有大肠杆菌(原核)体系、酵母(真核)体系、和哺乳动物(真核)细胞表达系统,不同的表达体系具有不同的特点和适用范围,国际一线单抗企业一般同时拥有成熟的原核和真核表达体系。
总的来说,如果单抗药物包含Fc片段或需要翻译后修饰则必须使用哺乳动物细胞(如Herceptin、Rituxin、Erbitiux的生产),如果不需要Fc段,可使用细菌和酵母体系以降低成本。目前全球哺乳动物细胞、大肠杆菌和酵母的发酵量分别占到总量的50%、33%和17%。




 加载中...
加载中...