教学目的:
1. 掌握微生物的能量代谢和生物氧化类型。
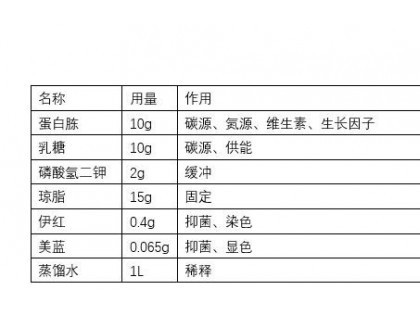
2. 掌握微生物对碳水化合物、蛋白质、脂肪的分解过程及分解代谢途径的特点和生理意义。
3. 掌握微生物各种发酵途径及其在发酵工业中的应用。
4. 理解次生代谢与次生代谢产物及调控的原理。
教学重点与难点:
1. 呼吸、无氧呼吸和发酵的异同点;发酵的类型;次生代谢与次生代谢产物及调控。
2. 微生物对碳水化合物、蛋白质、脂肪的分解过程及分解代谢途径的特点和生理意义。
教学课时:5学时
教学方法:多媒体教学
教学内容:(参考:何国庆,贾英民主编,《食品微生物学》. 北京:中国农业大学出版社,2002. 第三章)
1. 微生物的代谢
微生物同其他生物一样都是具有生命的,新陈代谢作用贯穿于它们生命活动的始终,新陈代谢作用包括合成代谢(同化作用)和分解代谢(异化作用)。微生物细胞直接同生活环境接触,微生物不停地从外界环境吸收适当的营养物质,在细胞内合成新的细胞物质和贮藏物质,并储存能量,即同化作用,这是其生长、发育的物质基础;同时,又把衰老的细胞物质和从外界吸收的营养物质进行分解变成简单物质,并产生一些中间产物作为合成细胞物质的基础原料,最终将不能利用的废物排出体外,一部分能量以热量的形式散发,这便是异化作用。在上述物质代谢的过程中伴随着能量代谢的进行,在物质的分解过程中,伴随着能量的释放,这些能量一部分以热的形式散失,一部分以高能磷酸键的形式贮存在三磷酸腺苷(ATP)中,这些能量主要用于维持微生物的生理活动或供合成代谢需要。
根据微生物代谢过程中产生的代谢产物在微生物体内的作用不同,又可将代谢分成初级代谢与次级代谢两种类型。初级代谢是指能使营养物质转换成细胞结构物质、维持微生物正常生命活动的生理活性物质或能量的代谢。初级代谢的产物成为初级代谢产物。次级代谢是指某些微生物进行的非细胞结构物质和维持其正常生命活动的非必须物质的代谢。如一些微生物积累发酵产物的代谢过程(抗生素、毒素、色素等)。
微生物的代谢作用是由微生物体内一系列有一定次序的、连续性的生物化学反应所组成,这些生化反应在生物体内可以在常温、常压和pH中性条件下极其迅速地进行,这是由于生物体内存在着多种多样的酶和酶系,绝大多数的生化反应是在特定酶催化下进行的。
同化作用和异化作用它们两者既是矛盾的,又是统一的,微生物同其它生物一样,新陈代谢作用是它最基本的生命过程,也是其它一切生命现象的基础。
1.1 微生物的能量代谢(重点理解其氧化类型的区别)
微生物在生命活动中需要能量,它主要是通过生物氧化而获得能量。所谓生物氧化就是指细胞内一切代谢物所进行的氧化作用。它们在氧化过程中能产生大量的能量,分段释放,并以高能磷酸键形式储藏在ATP分子内,供需要时用。
1.1.1微生物的呼吸(生物氧化)类型
根据在底物进行氧化时,脱下的氢和电子受体的不同,微生物的呼吸可以分为三个类型,即:好氧呼吸、厌氧呼吸、发酵。
⑴ 好氧呼吸(aerobic respiration)
以分子氧作为最终电子受体的生物氧化过程,称为好氧呼吸。许多异养微生物在有氧条件下,以有机物作为呼吸底物,通过呼吸而获得能量。以葡萄糖为例,通过EMP途径和TCA循环被彻底氧化成二氧化碳和水,生成38个ATP,化学反应式为:
C6H12O6+6O2+38ADP+38Pi→6CO2+6H2O+38ATP
⑵ 厌氧呼吸(anaerobic respiration)
以无机氧化物作为最终电子受体的生物氧化过程,称为厌氧呼吸。能起这种作用的化合物有硫酸盐、硝酸盐和碳酸盐。这是少数微生物的呼吸过程。例如脱氮小球菌利用葡萄糖氧化成二氧化碳和水,而把硝酸盐还原成亚硝酸盐(故称反硝化作用),反应式如下:
C6H12O6+12NO3– → 6CO2+6H2O+12NO2–+429000卡
⑶ 发酵作用(fermentation)
如果电子供体是有机化合物,而最终电子受体也是有机化合物的生物氧化过程称为发酵作用。在发酵过程中,有机物既是被氧化了基质,又是最终的电子受体,但是由于氧化不彻底,所以产能比较少。酵母菌利用葡萄糖进行酒精发酵,只释放2.26×105J热量,其中只有9.6×104J贮存于ATP中,其余又以热的形式丧失,反应式如下:
C6H12O6+2ADP+2Pi → 2C2H5OH+2CO2+2ATP
1.1.2 生物氧化链
微生物从呼吸底物脱下的氢和电子向最终电子受体的传递过程中,要经过一系列的中间传递体,并有顺序地进行,它们相互“连控”如同链条一样,故称为呼吸链(生物氧化链)。它主要由脱氢酶、辅酶Q和细胞色素等组分组成。它主要存在于真核生物的线粒体中;在原核生物中,则和细胞膜、中间体结合在一起。它的功能是传递氢和电子,同时将电子传递过程中释放的能量合成ATP。
1.1.3 ATP的产生
生物氧化的结果不仅使许多还原型辅酶Ⅰ得到了再生,而且更重要的是为生物体的生命活动获得了能量。ATP的产生就是电子从起始的电子供体经过呼吸链至最终电子受体的结果。
ATP是生物体内能量的主要传递者。当微生物获得能量后,都是先将它们转换成ATP。当需要能量时,ATP分子上的高能键水解,重新释放出能量。这些能量在体内很好地和起催化作用的酶产生偶然作用,既可利用,又可重新贮存。在pH为7.0的情况下,ATP的自由能变化△G是-3×104J,这种分子既比较稳定,又能比较容易引起反应,是微生物体内理想的能量传递者。因此ATP对于微生物的生命活动具有重大的意义。
利用光能合成ATP的反应,称为光合磷酸化。利用生物氧化过程中释放的能量,合成ATP的反应,称为氧化磷酸化,生物体内氧化磷酸化是普遍存在的,有机物降解反应和生成物合成反应通过氧化还原而偶联起来,使能量得到产生、保存和释放。
微生物通过氧化磷酸化生成ATP的方式有两种:
⑴ 底物水平磷酸化
在底物水平磷酸化中,异化作用的中间产物的高能磷酸转移给ADP,形成ATP,如下述反应:
磷酸烯醇丙酮酸 + ADP ————→ 丙酮酸 + ATP
⑵ 电子传递磷酸化
在电子传递磷酸化中,通过呼吸链传递电子,将氧化过程中释放的能量和ADP的磷酸化偶联起来,形成ATP。一个NAD分子,通过呼吸链进行氧化,可以产生3个ATP分子。它分别在三个位置,各产生一个ATP。第1个ATP大约在辅酶Ⅰ和黄素蛋白之间;第2个ATP大约在细胞色素b和c1之间;在第3个ATP大约在细胞色素c和a之间。如图3-2所示。
⑶ 光合磷酸化的能量转换
光能是一种辐射能,它能被微生物直接利用,只有当光能被光合生物的光合色素吸收并转变成化学能——ATP 以后,才能用于微生物的代谢或其它生理活动。可见光能转换是光合生物获得能量的一种主要方式。
光合色素是光合生物所特有的物质,它在光能转换过程中起着重要作用,光合色素由主要色素和辅助色素构成,主要色素是叶绿素或细菌叶绿素;辅助色素是类胡萝卜素和藻胆素。光合色素存在于一定的细胞器或细胞结构中,主要色素在它存在的部位里构成光反应中心,并能吸收光和捕捉光能,使自己处于激发态而逐出电子。辅助色素在细胞内只能捕捉光能并将捕捉到的光能传递给主要色素。光反应中心的叶绿素通过吸收光能逐出电子而使自己处于氧化态,逐出的电子通过铁氧还蛋白、泛醌、细胞色素b与细胞色素c组成的电子传递链再返回叶绿素本身,使叶绿素分子回复到原来的状态,在电子传递过程中产生能量转化(光能)—→化学能)。这种由光能引起叶绿素分子逐出电子,并通过电子传递来产生ATP的方式称为光合磷酸化。
1.2 微生物的分解代谢
地球上最丰富的有机物是纤维素,半纤维素,淀粉等糖类物质,自然界中微生物赖以生存的主要也是糖类物质,人们培养微生物,进行食品加工和工业发酵等也是以糖类物质为主要的碳源和能源物质。因此,微生物的糖代谢是微生物代谢的一个重要方面,掌握这方面的知识,对于认识自然界不同的微生物类群,以及搞好微生物的培养利用都是重要的基础知识。
1.2.1 微生物糖代谢的途径(重点比较学习,注意关键步骤和关键酶)
微生物糖代谢的主要途径有:EMP途径(Embden-Meverhef-Parnus Pathway),HMP途径(Hexose-Mono-Phosphate Pathway),E.D途径(Entner-Doudorof Pathway),Pk途径(Phosphoketolase pathway),等四种。
⑴ EMP途径
EMP途径也称已糖双磷酸降解途径或糖酵解途径。这个途径的特点是当葡萄糖转化成1.6-二磷酸果糖后,在果糖二磷酸醛缩酶作用下,裂解为两 个3C化合物,再由此转化为2分子丙酮酸。EMP途径的过程由以下10个连续反应组成:
(己糖激酶)
1) 葡萄糖 + ATP ——————→ 6-磷酸葡萄糖 + ADP
(磷酸己糖异构酶)
2) 6-磷酸葡萄糖 ——————→ 6-磷酸果糖
(磷酸己糖激酶)
3) 6-磷酸果糖 + ATP ——————→ 1,6- 磷酸果糖 + ADP
(醛缩酶)
4) 1,6-二磷酸果糖 ——————→ 磷酸二羟丙酮 + 3-磷酸甘油醛
(磷酸丙糖异构酶)
5) 磷酸二羟丙酮 ——————→ 3-磷酸甘油醛
(3-磷酸甘油醛脱氢酶)
6) 3-磷酸甘油醛 + NAD + H3PO4 ——→ 1,3-二磷酸甘油酸 + NADH
(3-磷酸甘油酸激酶)
7) 1,3-二磷酸甘油酸 + ADP ————————→ 3- 磷酸甘油酸 + ATP
(磷酸甘油酸变位酶)
8) 3-磷酸甘油酸 ————————→ 2-磷酸甘油酸
(烯醇化酶)
9) 2-磷酸甘油酸 ————————→ 磷酸烯醇式丙酮酸 + H2O
(丙酮酸激酶)
10) 磷酸烯醇式丙酮酸 +ADP ——————→ 丙酮酸 + ATP
总反应式为: C6H12O6+2NAD+2(ADP+Pi)→2CH3COCOOH+2ATP+2NADH2
EMP 途径的关键酶是磷酸已糖激酶和果糖二磷酸醛缩酶,它开始时消耗 ATP,后来又产生ATP,总计起来,每分子葡萄糖通过EMP途径净合成2分子ATP,产能水平较低。
EMP 途径是生物体内6-磷酸葡萄糖转变为丙酮酸的最普遍的反应过程,许多微生物都具有EMP途径。但EMP途径往往是和HMP途径同时存在于同一种微生物中,以EMP途径作为一唯一降解途径的微生物极少,只有在含有牛肉汁酵母膏复杂培养基上生长的同型乳酸细菌可以利用EMP作为唯一降解途径。EMP途径的生理作用主要是为微生物代谢提供能量(即ATP),还原剂(即NADH2)及代谢的中间产物如丙酮酸等。
在EMP途径的反应过程中所生成的NADH2不能积累,必须被重新氧化为NAD后,才能保证继续不断地推动全部反应的进行。NADH2重新氧化的方式,因不同的微生物和不同的条件而异。厌氧微生物及兼厌氧性微生物在无氧条件下,NADH2的受氢体可以是丙酮酸,如乳酸细菌所进行的乳酸发酵,也可以是丙酮酸的降解产物——乙醛,如酵母的酒精发酵等。好氧性微生物和在有氧条件下的兼厌氧性微生物经EMP途径产生的丙酮酸进一步通过三羧酸循环,被彻底氧化,生成CO2,氧化过程中脱下的氢和电子经电子传递链生成H2O和大量ATP。
三羧酸循环,简称TCA环(Tricarboxylic Acid Cycle)。TCA环的总反应式为:
CH3COSCoA+2O2+12(ADP+Pi)→2 CO2+ H2O +12ATP+ CoA
TCA 环产生能量的水平是很高的,每氧化一分子乙酰CoA,可产生12分子ATP。
葡萄糖经EMP途径和TCA 环彻底氧化成CO2和H2O的全部过程为:
① C6H12O6+2NAD+2(ADP+Pi)→2 CH3COCOOH +2ATP+2 NADH2
2 NADH2+O2+6(ADP+Pi)→2NAD+2 H2O +6ATP
② 2 CH3COCOOH+2NAD+2 CoA→2 CH3COSCoA +2 CO2+2 NADH2
2 NADH2+O2+6(ADP+Pi)→2NAD+2 H2O +6ATP
③ 2 H3COSCoA +4O2+24(ADP+Pi)→4 CO2+2 H2O +24ATP+2 CoA
总反应式: C6H12O6+6O2+38(ADP+Pi)→6 CO2+6 H2O +38ATP
TCA循环的关键酶是柠檬酸合成酶,它催化草酰乙酰与乙酰CoA合成柠檬酸的反应。很多微生物中都存在这条循环途径,它除了产生大量能量,作为微生物生命活动的主要能量来源以外,还有许多生理功能。特别是循环中的某些中间代谢产物是一些重要的细胞物质,如各种氨基酸、嘌呤、嘧啶及脂类等生物合成前体物,例如乙酰CoA是脂肪酸合成的起始物质;α-酮成二酸可转化为谷氨酸,草酰乙酸可转化为天门冬氨酸,而且上述这些氨基酸还可转变为其他氨基酸,并参与蛋白质的生物合成。另外,TCA环不仅是糖有氧降解的主要途径,也是脂、蛋白质降解的必经途径,例如脂肪酸经β-氧化途径,变成乙酰CoA可进入TCA环彻底氧化成CO2和H2O;又如丙氨酸,天门冬氨酸,谷氨酸等经脱氨基作用后,可分别形成丙酮酸,草酰乙酸,α-酮戊二酸等,它们都可进入TCA环被彻底氧化。因此,TCA环实际上是微生物细胞内各类物质的合成和分解代谢的中心枢纽。
由于EMP途径和TCA环研究得比较清楚,在发酵工业中得到了广泛地应用。用一种方法来阻止某一阶段的进行,就必然积累某些中间产物。根据这一原理,工业上已筛选出一些优良菌株,进行工业发酵,生产柠檬酸,异柠檬酸,α-酮戊二酸,苹果酸等。例如利用黑曲霉生产柠檬酸时,由于菌体内顺乌头酸水解酶的活力特别低,使柠檬酸大量积累。
⑵ HMP途径
也称已糖单磷降解途径或磷酸戊糖循环。这个途径的特点是当葡萄糖经一次磷酸化脱氢生成6-磷酸葡萄糖酸后,在6-磷酸葡萄糖酸脱酶作用下,再次脱氢降解为1分子CO2和1分子磷酸戊糖。磷酸戊糖的进一步代谢较复杂,由3分子磷酸已糖经脱氢脱羧生成的3分子磷酸戊糖, 3分子磷酸戊糖之间,在转酮酶和转醛酶的作用下,又生成2分子磷酸己糖和一分子磷酸丙糖,磷酸丙糖再经EMP途径的后半部反应转为丙酮酸,这个反应过程称为HMP途径。反应步骤可分为十一步反应:(略)
完全HMP途径的总反应式为:
6-磷酸葡萄糖 + 7H2O + 12NADP ——→ 6CO2+ 12NADPH2+ H3PO4
不完全HMP途径反应到(9)步反应为止。所生成的3-磷酸甘油醛经过EMP途径的后半部分,转化成丙酮酸。不完全HMP途径的总反应式为:
6-磷酸葡萄糖 + 7H2O + 12NADP → CH3COCOOH + 3CO2+ 6NADPH2+ ATP
HMP 途径的关键酶系是6-磷酸葡萄糖酸脱氢酶和转酮 一 转醛酶系,其中6-磷酸葡萄糖酸脱氢酶催化磷酸已糖酸的脱氢脱羧,而转酮 一 转醛酶系则作用于三碳糖、四碳糖、五碳糖、六碳糖及七碳糖的相互转化。
HMP途径的另一特点是只有NADP参与反应。在有氧条件下,HMP途径所产生的NADPH2在转氢酶的作用下,可将氢转给NAD,形成NADH2,经呼吸链,将电子和氢交给分子态氧形成水,并由电子传递磷酸化作用形成ATP。但是一般认为HMP途径不是主要的产能途径,而是为细胞的生物合成提供供氢体(NADPH2)。另外,HMP途径还为细胞生物合成提供大量的C3、C4、C5、C6和C7等前体物质,特别是磷酸戊糖,它是合成核酸,某些辅酶以及合成组氨酸,芳香族氨酸,对氨基苯甲酸等化合物的重要底物。此外,HMP途径与化能自养菌和光合细菌的碳代谢有密切联系。因此,HMP途径的生理功能是多方面的,在微生物代谢中占有重要的地位。
HMP途径普遍存在于微生物细胞中,通常是和EMP途径同时存在一种微生物中。能以HMP途径作为唯一降解途径的微生物,目前发现的只有亚氧化醋酸杆菌(Acetobacter suboxydans)。
⑶ ED途径
也称2-酮-3-脱氧-6-磷酸葡萄糖酸途径,在醛缩酶(KDPGaldolase)的作用下,裂解为丙酮酸和3-磷酸甘油醛,3-磷酸甘油醛再经EMP途径的后半部反应转化为丙酮酸。
ED途径的关键酶系是6-磷酸葡萄糖脱水酶和2-酮-3-脱氧-6-磷酸葡萄糖酸醛缩酶。其中6-磷酸葡萄糖酸脱水导致2-酮-3-脱氧-6-磷酸葡萄糖酸的生长,而2-酮-3-脱氧-6-磷酸葡萄糖酸醛缩酶则催化2-酮-3-脱氧-6-磷酸葡萄糖酸裂解为丙酮酸和3-磷酸甘油醛。
ED途径是糖类的一个厌氧降解途径,它在细菌中,特别是革兰氏阴性细菌中分布很广,在好氧菌中分布不普遍。例如嗜糖假单胞杆菌(Pseudomonas saccharophila),发酵假单胞菌(Zymomonas mobilos)以及铜绿色假单胞杆菌(Pseudomonas aeruginasa)等中都具有ED途径。这个途径多数情况下是与HMP途径同时存在于一种微生物中,但也可以独立存在于某些细菌中。
总反应式为:C6H12O6+
ADP+Pi+NADP+NAD→2 CH3COCOOH +ATP+ NADPH2+ NADH2
⑷ PK途径
也称磷酸解酮酶途径。在微生物降解已糖的过程中,除了EMP、HMP和E.D途径外,还有一条途径即磷酸解酮酶途径(Phosphoketolase Pathway)为少数细菌所独有。磷酸解酮酶由两种,一种是戊糖磷酸解酮酶,一种是己糖磷酸解酮酶;有些异型乳酸发酵的微生物,如明串珠菌属(Leuconostoc)和乳杆菌属(Lactobacillus) 中的肠膜明串球菌(Leuconostoc mesenteulides),短乳酸杆菌(Lactobacillus brevie),甘露乳酸杆菌(Lactobacillus manitopoeum)等,由于没有转酮-转醛酶系,而具有戊糖磷酸解酮酶,因此就不能通过HMP途径进行异型乳酸发酵,而是通过戊糖磷酸解酮酶途径进行的。
这个途径的特点是降解1分子葡萄糖只产生1分子ATP,相当于EMP途径的一半,另一特点是几乎产生等量的乳酸,乙醇和CO2。总反应式为:
C6H12O6+ADP+Pi→CH3CHOHCOOH+CH3CH2OH+CO2+ATP
戊糖磷酸解酮途径的关键酶系是磷酸木酮糖解酮酶,它催化5-磷酸木酮糖裂解为3-磷酸甘油醛和乙酰磷酸的反应。
1.2.2 多糖的分解(详细了解,因为牵涉到微生物在食品工业中的应用)
多糖分解的种类很多,如淀粉、纤维素、果胶质的分解。
⑴ 淀粉的分解
淀粉是多种微生物用作碳源的原料。它是葡萄糖的多聚物,有直链淀粉和支链淀粉之分。
微生物对淀粉的分解是由微生物分泌的淀粉酶催化进行的。淀粉酶是水解淀粉糖苷键一类酶的总称,它的种类有以下几种:
① 液化型淀粉酶(又称α-淀粉酶) 这种酶可以任意分解淀粉的α-1,4糖苷键,而不能分解α-1,6糖苷键。淀粉经该酶作用以后,粘度很快下降,液化后变为糊精,最终产物为糊精、麦芽糖和少量葡萄糖。由于这种酶能使淀粉表现为液化,淀粉粘度急速下降,故称液化酶,又由于生成的麦芽糖在光学上是α型,所以又称为α-淀粉酶。
产生α-淀粉酶的微生物很多,细菌、霉菌、放线菌中的许多种都能产生。
② 糖化型淀粉酶 这类酶又可细分为好几种,其共同特点将淀粉水解为麦芽糖或葡萄糖,故称为糖化型淀粉酶。
β-淀粉酶(淀粉1,4-麦芽糖苷酶):此酶作用方式是从淀粉分子的非还原性末端开始,逐次分解。分解物以麦芽糖为单体,但不能作用于也不能越过α-1,6糖苷键,这样分解到最后,仍会剩下较大分子的极限糊精。由于生成的麦芽糖,在光学是上β型,所以称为β-淀粉酶。
糖化酶(淀粉1,4、1,6-葡萄糖苷酶):此酶对α-1,4-糖苷键能作用,对α-1,6-糖苷键也能分解,所以最终产物几乎全是葡萄糖。常用于生产糖化酶的菌种有根霉、曲霉等。
异淀粉酶(淀粉1,6-糊精酶):此酶可以分解淀粉中的α-1,6-糖苷键,生成较短的直链淀粉。异淀粉酶用于水解由α-淀粉酶产生的极限糊精和由β-淀粉酶产生的极限糊精。
异淀粉酶存在于产气气杆菌、中间埃希氏杆菌、软链球菌、链霉菌等。
微生物产生的淀粉酶广泛用于粮食加工、食品工业、发酵、纺织、医药、轻工、化工等行业。
⑵ 纤维素的分解
纤维素的葡萄糖由β-1,4糖苷键组成的大分子化合物。它广泛存在于自然界,是植物细胞壁的主要组成成分。人和动物均不能消化纤维素。但是很多微生物,例如木霉、青霉、某些放线菌和细菌均能分解利用纤维素,原因是它们能产生纤维素酶。
纤维素酶是一类纤维素水解酶的总称。它由c1酶、cx酶和解成纤维二糖,再经过β-葡萄糖苷酶作用,最终变为葡萄糖,其水解过程如下:
C 1酶 Cx1 Cx2酶 纤维二糖酶
天然纤维素 ——→ 水合纤维素分子 ——→ 纤维二糖 ——→ 葡萄糖
生产纤维素酶的菌种常有绿色木霉、康氏木霉、某些放线菌和细菌。我国采用绿色木霉、木素木霉为菌种,进行了研究、试制。
纤维素酶在为开辟食品及发酵工业原料新来源,提高饲料的营养价值,综合利用农村的农付产品方面将会起着积极的作用,具有重要的经济意义。
⑶ 果胶质的分解
果胶是植物细胞的间隙物质,使邻近的细胞壁相连,是半乳糖醛酸以α-1,4糖苷键结合成直链状分子化合物。其羧基大部分形成甲基酯,而不含甲基酯的称为果胶酸。
果胶在浆果中最丰富。它的一个重要特点是在酸和糖存在下,可以形成果冻。食品厂利用这一性质来制造果浆、果冻等食品;但对果汁加工、葡萄酒生产引起榨汁困难。
果胶酶含有不同的酶系,在果胶分解中起着不同的作用。主要有果胶酯酶和半乳糖醛酸酶两种,引起的反应式如下:
果胶酯酶 聚半乳糖醛酸酶
果胶 ——————→ 甲醇+果胶酸 ————————→ 半乳糖醛酸
果胶酶广泛存在于植物、霉菌、细菌和酵母中。其中以霉菌产的果胶酶产量高,澄清果汁力强,因此工业上常用的菌种几乎都是霉菌,例如文氏曲霉、黑曲霉等。果胶酶大多属于诱导酶,故生产时必须添加含果胶的物质,才会提高产量。
1.2.3 蛋白质的分解
⑴ 蛋白质的分解
蛋白质是由氨基酸组成的分子巨大、结构复杂的化合物。它们不能直接进入细胞。微生物利用蛋白质,首先分泌蛋白酶至体外,将其分解为大小不等的多肽或氨基酸等小分子化合物后再进入细胞。通式如下:
蛋白酶
蛋白质 ——→ 多肽、氨基酸
产生蛋白酶的菌种很多,细菌、放线菌、霉菌等中均有。不同的菌种可以产生不同的蛋白酶,例如黑曲霉主要生产酸性蛋白酶。短小芽孢杆菌用于生产碱性蛋白酶。不同的菌种也可生产功能相同的蛋白酶,同一个菌种也可产生多种性质不同的蛋白酶。
⑵ 氨基酸的分解(可稍略)
微生物对氨基酸的分解,主要是脱氨作用和脱羧基作用。
1)脱氨作用 脱氨方式随微生物种类、氨基酸种类以及环境条件的不同,也不一样。主要有以下几种:
① 氧化脱氨。在酶催化下,氨基酸在氧化脱氢的同时释放游离氨,这一过程即氧化脱氨。这种脱氨方式须在有氧气条件下进行。专性厌氧菌不能进行氧化脱氨。微生物催化氧化脱氨的酶有两类:一类是氨基氧化酶,以FAD或FMN为辅基;另一类是氨基酸脱氢酶,以NAD或NADP作为氢的载体,交给分子态氧。反应式如下:
2R-CHNH2-COOH + O2 ——→ 2R-CO-COOH + 2NH3
实际上,这一反应是由脱氢和加水两个反应步骤完成的:
氧化酶
R-CHNH2 -COOH + O2 ——→ R-C=NH-COOH + H2 O2
FAD
非酶促反应
R-C=NH-COOH + H2O2 ——→ R-CO-COOH + NH3
② 还原脱氨。还原脱氨在无氧条件下进行,生成饱和脂肪酸。能进行还原脱氨的微生物是专性厌氧菌和兼性厌氧菌。腐败的蛋白质中常分离到饱和脂肪酸便是由相应的氨基酸生成。如大肠杆菌可使甘氨酸还原脱氨成乙酸,反应式如下:
NADH2 NAD
③ 水解脱氨。不同氨基酸经水解脱氨生成不同的产物。同种氨基酸水解之后也可形成不同的产物,反应通式如下:
水解酶
R-CHNH2 -COOH + H2O ——→ R-CHOH-COOH + NH3
有些细菌可以水解色氨酸生成吲哚,吲哚可以与二甲基氨基苯甲醛反应生成红色的玫瑰吲哚,因此可根据细菌能否分解色氨酸产生吲哚来鉴定菌种
④ 减饱和脱氨(直接脱氨)。氨基酸在脱氨的同时,其α.β键减饱和,结果生成不饱和酸。例如天门冬氨酸减饱和脱氨生成延胡索酸,反应式如下:
天门冬氨酸裂解酶
HOO-CH2-CHNH2-COOH ——————→ HOOC-CH=CH-COOH + NH3
⑤ 氧化—还原偶联脱氨(Stickland 反应)。Stickland发现在某些梭菌中存在由两种氨基酸参与的脱氨基反应,一种氨基酸被氧化,在脱氨的同时脱下氢和电子,同时另一种氨基酸被还原,得到氢和电子的同时脱下氨基。丙氨酸和甘氨酸的偶联反应如下:
CH3CHNH2COOH ——→ CH2NH2COOH
NAD
NADH
CH3COCOOH + NH3 ——→ CH3COOH + NH3
这种偶联反应并不是在任意两种氨基酸之间就能发生的,有些氨基酸优先作为供氢体,有些优先作为受氢体(见表5-1)。
表5-1 在进行Stickland 反应时的供氢体和受氢体的氨基酸对
供 氢 体 受 氢 体
L-丙氨酸 L-丙氨酸
L-亮氨酸 L-亮氨酸
L-异亮氨酸 L-异亮氨酸
L-缬氨酸 L-缬氨酸
L-组氨酸 L-组氨酸
L-苯丙氨酸 L-苯丙氨酸
L-丝氨酸 L-丝氨酸
2) 脱羧作用 氨基酸脱羧作用常见于许多腐败细菌和真菌中。不同的氨基酸由相应的氨基酸脱羧酶催化脱羧,生成减少一个碳原子的胺和二氧化碳,通式如下:
氨基酸脱梭酶
R-CHNH2-COOH ————→ R-CH2-NH2 + CO2
脱羧酶具有高度专一性,需要磷酸吡哆醛为辅酶,大多数是诱导酶。一元氨基酸脱羧后变成一元胺;二元氨基酸脱羧后变成二元胺。这类物质统称为尸碱,有毒性。肉类蛋白质腐败后常生成二元胺,故不能食用。例如赖氨酸脱羧后变成尸胺,反应式如下:
赖氨酸脱梭酶
H2N(CH2)4CHNH2COOH ——→ H2N(CH2)4CH2NH2+CO2
1.2.4 脂肪和脂肪酸的分解
⑴ 脂肪的分解
脂肪是脂肪酸的甘油三酯。在脂肪酶作用下,可水解生成甘油和脂肪酸。
脂肪酶成分较为复杂,作用对象也不完全一样。不同的微生物产生的脂肪酶作用也不一样。能产生脂肪酶的微生物很多,有根霉、圆柱形假丝酵母、小放线菌、白地霉等。
脂肪酶目前主要用于油脂工业、食品工业、纺织工业上。常用作消化剂、乳品增香、制造脂肪酸、绢丝的脱脂等。
⑵ 脂肪酸的分解
微生物分解脂肪酸主要是通过β-氧化途径。β-氧化是由于脂肪酸氧化断裂发生在β-碳原子上而得名。在氧化过程中,能产生大量的能量,最终产物是乙酰辅酶A。而乙酰辅酶A 是进入三羧酸循环的基本分子单元。
1.3 微生物发酵的代谢途径
由于微生物种类繁多,能在不同条件下对不同物质或对基本相同的物质进行不同的发酵。而不同微生物对不同物质发酵时可以得到不同的产物,不同的微生物对同一种物质进行发酵,或同一种微生物在不同条件下进行发酵都可得到不同的产物,这些都取决于微生物本身的代谢特点和发酵条件,现将食品工业中常见的微生物及其发酵途径介绍如下:
1.3.1 醋酸发酵
参与醋酸发酵的微生物主要是细菌,统称为醋酸细菌。它们之中既有好氧性的醋酸细菌,例如纹膜醋酸杆菌(Acetobacter aceti),氧化醋酸杆菌(Acetobacter oxydans),巴氏醋
好氧性的醋酸细菌进行的是好氧性的醋酸发酵,在有氧条件下,能将乙醇直接氧化为醋酸,是醋酸细菌的好氧性呼吸,其氧化过程是一个脱氢加水的过程。
脱下的氢最后经呼吸链和氧结合形成水,并放出能量:
4H+O2 ——→ 2 H2O +117千卡
总反应式为: CH3CH2OH+O2 ——→ CH3COOH+ H2O +117千卡
厌氧性的醋酸细菌进行的是厌氧性的醋酸发酵,其中热醋酸梭菌能通过EMP途径发酵葡萄糖,产生3M醋酸。研究证明该菌只有丙酮酸脱羧酶和COM,能利用CO2作为受氢体生成乙酸,总反应式:
C6H12O6 + 4(ADP+Pi)→3CH3COOH+4ATP
好氧性的醋酸发酵是制醋工业的基础。制醋原料或酒精接种醋酸细菌后,即可发酵生成醋酸发酵液供食用,醋酸发酵液还可以经提纯制成一种重要的化工原料——冰醋酸。厌氧性的醋酸发酵是我国用于酿造糖醋的主要途径。
1.3.2 柠檬酸发酵
关于柠檬酸发酵途径曾有多种论点,但目前大多数学者认为柠檬酸并非单纯由TCA循环所积累,而是由葡萄糖经EMP途径形成丙酮酸,再由两分子丙酮酸之间发生羧基转移,形成草酰乙酸和乙酰CoA, 草酰乙酸和乙酰CoA再缩和成柠檬酸,其反应途径如图5-7。
CO2 柠檬酸
丙酮酸 ——————→ 乙酰辅酶A
图5-7 柠檬酸发酵代谢途径
能够累积柠檬酸的霉菌以曲霉属(Aspergillus),青霉属(Penicillium)和桔霉属(Citromyces)为主。其中以黑曲霉(Asp. niger)、米曲霉(Asp.oryzae),灰绿青霉(Pen. glaucum),淡黄青霉(Pen.luteum),光桔霉(Citromyces glaber)等产酸量最高。
柠檬酸发酵广泛被用于制造柠檬酸盐、香精、饮料、糖果、发泡缓冲剂等,在食品工业中起重要的作用。
1.3.3酒精发酵
酒精发酵是酿酒工业的基础,它与酿造白酒,果酒,啤酒以及酒精的生产等有密切关系。进行酒精发酵的微生物主要是酵母菌,如啤酒酵母(Saccharomyces cerevisiae)等,此外还有少数细菌如发酵单胞菌(Zymononas mobilis),嗜糖假单胞菌(Pseudomonas Saccharophila),解淀粉欧文氏菌(Eruinia amylovora)等也能进行酒精发酵。
酵母菌在无氧条件下,将葡萄糖经EMP途径分解为2分子丙酮酸,然后在酒精发酵的关键酶——丙酮酸脱羧酶的作用下脱羧生成乙醛和CO2,最后乙醛被还原为乙醇。
总反应式:
C6 H12 O6+2ADP+2Pi ————→ 2 CH3CH2 OH+2CO2+2ATP
酒精发酵是酵母菌正常的发酵形式,又称第一型发酵,如果改变正常的发酵条件,可使酵母进行第二型和第三型发酵而产生甘油。第二型发酵是在有亚硫酸氢钠的存在的情况下发生的。亚硫酸氢钠和乙醛起加成作用,生成难溶的结晶状亚硫酸氢钠加成物--磺化羟乙醛.
由于乙醛和亚硫氢钠发生了加成作用,致使乙醛不能作为受氢体,而迫使磷酸二羟丙酮代替乙醛作为受氢体生成a-磷酸甘油,α-磷酸甘油在α-磷酸甘油磷酸酯酶催化下被水解,除去磷酸而生成甘油:
第三型发酵是在碱性条件下进行的,碱性条件可促使乙醛不能作为正常的受氢体,而是两分子乙醛之间发生歧化反应,即相互进行氧化还原反应,一分子乙醛被氧化成乙酸,另一分子乙醛被还原为乙醇:
CH3CHO + CH3CHO ——→CH3COOH + CH3CH2OH
这样又迫使磷酸二羟丙酮作为受氢体而最终形成甘油。
总反应式为: 2C6H12O6→2甘油+乙酸+乙醇+2 CO2
由此可以看出,酵母菌的第二型和第三型发酵过程中,都不产生能量,因此只能在非生长情况下进行。如用此途径生产甘油,必需在第三型发酵液中不断地加入碳酸钠以维持其碱性,否则由于酵母菌产生酸而使发酵液pH 降低,这样就又恢复到正常的第一型发酵而不累积甘油。这说明酵母菌在不同条件下发酵结果是不同的,因而我们可以通过控制环境条件来利用微生物的代谢活动,有目的地生产有用的产品。
1.3.4 乳酸发酵
乳酸是细菌发酵最常见的最终产物,一些能够产生大量乳酸的细菌称为乳酸细菌。在乳酸发酵过程中,发酵产物中只有乳酸的称为同型乳酸发酵;发酵产物中除乳酸外,还有乙醇、乙酸及CO2等其它产物的,称为异型乳酸发酵。
⑴ 同型乳酸发酵
引起同型乳酸发酵的乳酸细菌,称为同型乳酸发酵菌,有双球菌属(Diplococcus)、链球菌属(Streptococcus)及乳酸杆菌属(Lactobacillus)等。其中工业发酵中最常用的菌种是乳酸杆菌属中的一些种类,如德氏乳酸杆菌(L.delhruckii)、保加利亚乳酸杆菌(L.bulgaricus)、干酪乳酸杆菌(L.casei)等。
同型乳酸发酵的基质主要是已糖,同型乳酸发酵菌发酵已糖是通过EMP途径产生乳酸的。其发酵过程是葡萄糖经EMP途径降解为丙酮酸后,不经脱羧,而是在乳酸脱氢酶的作用下,直接被还原为乳酸:
总反应式:C6 H12 O6 +2ADP+2Pi→2CH3CHOHCOOH+2ATP
⑵ 异型乳酸发酵
异型乳酸发酵基本都是通过磷酸解酮酶途径(即PK途径 )进行的。其中肠膜明串球菌(Leuconostos mesentewides)葡萄糖明串球菌(Leuconostoc dextranicum),短乳杆菌(Lactabacillus brevis)蕃茄乳酸杆菌(Lactobacillus lycopersici)等是通过戎糖解酮酶途径将1分子葡萄糖发酵产生1分子乳酸,1分子乙醇和1分子CO2,并且只产生1分子ATP。总反应式如下:
C6 H12 O6+ADP+Pi→CH3CHOHCOOH+CH3CH2OH+CO2+ATP
双*乳酸杆菌(Lactobacillus bifidus),两歧双歧乳酸菌(Bifidobacterium bifidus)等是通过已糖磷酸解酮酶途径将2分子葡萄糖发酵为2分子乳酸和3分子乙酸,并产生5分子ATP,总反应式为:
2 C6 H12 O6+5ADP+5Pi→2 CH3CHOHCOOH+3 CH3COOH+5ATP
乳酸发酵被广泛地应用于泡菜、酸菜、酸牛奶、乳酪以及青贮饲料中,由于乳酸细菌活动的结果,积累了乳酸,抑制其他微生物的发展,使蔬菜,牛奶及饲料得以保存。近代发酵工业多采用淀粉为原料,先经糖化,再接种乳酸细菌进行乳酸发酵生产纯乳酸。
1.4 微生物独特的合成代谢
所谓合成代谢,是指微生物利用能量将简单的无机或有机的小分子前体物质同化成高分子或细胞结构物质;但微生物合成代谢时,必须具备三个条件,那就是代谢能量、小分子前体物质和还原基, 只有具备了这三个基本条件,合成代谢才能进行。自养型微生物的合成代谢能力很强,它们利用无机物能够合成完全的自身物质;在食品工业,涉及最多的是化能异养型微生物,这些微生物所需要的代谢能量、小分子前体物质和还原基都是从复杂的有机物中获得,获得代谢能量、小分子前体物质和还原基的过程是微生物对吸收的营养物质的降解过程,所以,分解代谢和合成代谢是不能分开的,两者在生物体内是有条不紊的平衡过程。由于微生物蛋白质的合成和核酸的合成基本同一般生物的生化过程,限于篇幅,这里主要对微生物独特的肽聚糖的生物合成代谢途径加以阐述。
细胞壁肽聚糖的合成过程是一个极其复杂的过程,根据反应进行的部位不同,整个合成过程可分为在细胞质中、在细胞膜上和在细胞膜外三个阶段。
在细胞质中合成
1) 由葡萄糖合成N-乙酰葡萄糖胺和N-乙酰胞壁酸。
ATP ADP Gln Glu
葡萄糖 ——→ 6-磷酸-葡萄糖 ——→ 6-磷酸-果糖 ——→ 6-磷酸-葡萄糖胺
乙酰COA COA UTP PPi
磷酸烯醇丙酮酸 Pi
N-乙酰葡萄糖胺-UDP ————————————→ N-乙酰胞壁酸-UDP
NADPH2 NADP
2) 由N-乙酰胞壁酸合成“P”核苷酸 这一过程需要4步反应,它们都需要尿嘧啶二磷酸(UDP)作为糖的载体,另外还有合成D-丙氨酰胺-D-丙氨酸的两步反应,这些反应都可被环丝氨酸所抑制。
在细胞膜中合成
由“P”核苷酸合成肽聚糖亚单位的过程是在细胞膜上完成的,在细胞质内合成“P”核苷酸后,穿入细胞膜并进一步接上N-乙酰葡萄糖胺和甘氨酸五肽,即合成了肽聚糖亚单位。这个肽聚糖亚单位通过一个 类脂载体(十一异戊烯磷酸)携带到细胞膜外,进行肽聚糖合成。
由“P”核苷酸合成肽聚糖亚单位的过程总计有五步反应,
在细胞膜外的合成
被运送到细胞膜外的肽聚糖亚单位在必须有细胞壁残余(至少6~8个肽聚糖亚单位)作引物的条件下,肽聚糖亚单位与引物分子间先发生转糖基作用(transglycosylation)使多糖横向延伸一个双糖单位,然后,再通过转肽作用(transpeptidation)使两条多糖链间形成甘氨酸五肽而发生纵向交联反应。
青霉素可抑制转肽作用进行,其作用机制是:青霉素是肽聚糖亚单位五肽末端的D-丙氨酰胺-D-丙氨酸的类似物,两者竞争转肽酶的活性中心,从而竞争性抑制了肽聚糖的转肽作用,使得肽聚糖分子不能发生纵向交联反应, 肽聚糖不能形成细胞壁层。可见,青霉素的抑菌作用,只能是对处于活跃生长的细菌,对处于休眠阶段的细菌几乎无作用。
复习思考题:
1.何谓新陈代谢?试用图示说明合成代谢与分解代谢的相互关系。
2.什么是培养基?配制培养基的基本原则是什么?
3. 什么叫生物氧化?生物氧化与非生物氧化(燃烧)有何不同?
4. 什么叫发酵、有氧呼吸和无氧呼吸?试比较三者的异同。
5. 什么是呼吸链(电子传递链或生物氧化链)?它在生命活动中有何重要意义?
6. 什么叫底物水平磷酸化、氧化磷酸化和光合磷酸化?
7. 试比较光能型微生物和化能型微生物能量代谢的不同特点。
8. 微生物利用葡萄糖进行分解代谢的途径有哪些?每一代谢途径的特点和生理作用如何?
9. 试述TCA循环在微生物代谢和发酵生产中的重要性。
10. 简述微生物对淀粉、纤维素、果胶质的分解过程,各有哪些酶的参与?产酶微生物的种类如何?
11. 能够产生蛋白酶而分解蛋白质生成多肽和氨基酸的微生物有哪些?
12. 微生物分解氨基酸的脱氨方式有几种?试比较它们的不同之处。
13. 能够产生脂肪酶而分解脂肪生成脂肪酸和甘油的微生物有哪些?
14. 微生物通过哪种代谢途径分解脂肪酸?其最终产物是什么?
15. 什么是好氧性醋酸发酵和厌氧性醋酸发酵?二者在微生物种类和发酵途径上有何不同?各有哪些具体应用?
16. 能够积累柠檬酸的微生物有哪些?试用图示说明柠檬酸发酵途径。
17. 啤酒酵母利用糖类物质可进行哪几种类型的发酵作用?其发酵条件、发酵途径和最终产物有何不同?
18. 试述酵母菌酒精发酵和细菌酒精发酵的不同点。
19. 列表比较同型乳酸发酵和异型乳酸发酵的异同。
20.肽聚糖的生物合成分为哪几个阶段?试用简图表示每一阶段肽聚糖生物合成的途径。