, Linquan Bai , Zixin Deng
, Linquan Bai , Zixin Deng 链霉菌产生了一系列具有重要生物活性的次级代谢产物,Ⅰ型聚酮合酶催化形成的化合物(Ⅰ型聚酮类化合物)是一大类结构复杂、生物活性多样的天然产物。从化合物的结构上分析,该类化合物的结构类型涵盖了大环内酯类(红霉素)、安莎类(利福霉素)、多烯类(两性霉素)、聚醚类(盐霉素)等。在已发现的微生物药物中,该类化合物是重要的组成部分,且在市场上所销售的抗生素中占据重要的位置[1-2],引起了全世界研究者的关注。随着DNA测序技术的发展,越来越多的放线菌基因组被测序。在已测序的近千株放线菌基因组中,Ⅰ型聚酮类化合物的生物合成基因簇(Biosynthetic gene clusters,BGCs)数量众多,很可能是新药发现的重要资源[3-4]。现今人们生活方式的多样化与环境的恶化等因素,导致了新型疾病不断出现,一些病原菌的抗生素耐药问题也日益突出,迫切需要发掘具有新结构或者新生物活性的天然化合物[5],为新药研发提供更多的候选物质。
Streptomyces albus DSM 41398是盐霉素(聚醚类抗生素)的野生型产生菌,前期研究中,我们获得了该菌株的精确基因组信息,通过antiSMASH和RT-PCR分析显示,除了盐霉素的BGC外,该菌株染色体上还存在多个其他Ⅰ型聚酮类化合物的BGCs,其中有7个Ⅰ型聚酮类化合物的BGCs处于活跃表达的状态。为了减少盐霉素生物合成的前体分流,我们对上述7个基因簇进行系统缺失研究,得到一系列Ⅰ型聚酮类化合物BGCs的单独缺失突变菌株,分别为△PKS-1、△PKS-2、△PKS-3、△PKS-4、△PKS-5、△PKS-6和△PKS-9[6]。
在此基础之上,本研究通过系统比较野生型菌株和PKS中断突变菌株代谢产物的差异性,从S. albus DSM 41398的发酵产物中,发现了具有抗肿瘤活性的Ⅰ型聚酮类化合物放线吡喃酮(Actinopyranone)和洋橄榄菌素(Elaiophylin)。然后,根据其生物合成基因簇的信息及组成对其生物合成途径进行了分析与推导。本研究结果显示了S. albus DSM 41398具有产生多样化Ⅰ型聚酮类次生代谢产物的潜力,并且首次报道了放线吡喃酮的BGC,推导了其可能的生物合成途径,丰富了该类化合物生物合成途径的研究。此外,洋橄榄菌素BGC的发现与生物合成途径分析,也为其代谢工程优化及结构改造提供了较好的研究基础。
1 材料和方法1.1 材料1.1.1 菌株: 野生型Streptomyces albus DSM 41398,该菌株基因组序列的GenBank号为NZ_CP010519。突变株△PKS-1、△PKS-2、△PKS-3、△PKS-4、△PKS-5、△PKS-6和△PKS-9[6],本实验室保存。
1.1.2 主要生化试剂及仪器: 乙酸乙酯、石油醚、丙酮、三氯甲烷、甲醇购自国药集团化学试剂股份有限公司,乙腈购自安捷伦公司,氘代甲醇购自SIGMA公司;振荡摇床(ZQZY-70BS)为上海知楚仪器有限公司生产,高速离心机(SORVALL RC6+)为Thermo SCIENTIFIC公司生产,生物安全柜(HFsafe-1200)为上海力申科学仪器有限公司生产;采用Agilent 1262 series HPLC进行化合物的分析,化合物的精确分子量在Agilent G6530 Q-TOF上进行测定,核磁数据在Bruker Avance Ⅲ 600 MHz核磁共振仪上进行采集。
1.1.3 培养基: (1) 产孢培养基:ISP4培养基(购自BD-Difco公司)。(2)种子培养基(g/L):葡萄糖40、黄豆饼粉30、酵母粉10,碳酸钙2,pH 7.2。(3)发酵培养基(g/L):葡萄糖30.0、水解酪蛋白10.0、氯化钠2.0、氯化钾2.0、硫酸铵5.0、磷酸氢二钾0.2、七水硫酸镁0.1、氯化钙0.1,碳酸钙5.0,pH 7.2。
1.2 菌株的活化
将保存在20%甘油里的孢子涂在产孢培养基ISP4培养基上活化以便后续发酵。
1.3 菌株的发酵和发酵样品的处理发酵方法:分别将活化好的菌株接种到含有25 mL种子培养基的250 mL三角弹簧种子发酵瓶中,30 ℃、220 r/min摇床培养36 h后,按照10%的接种量转接到发酵培养基中,于220 r/min、30 ℃培养7 d。样品的处理方法:发酵完成后,发酵液用等体积的乙酸乙酯萃取3次。合并乙酸乙酯萃取液,进行减压干燥,再用1 mL甲醇将粗提物进行溶解,经过0.22 μm有机相微孔滤膜过滤后,供HPLC和LC-MS分析使用。大量发酵的方法:发酵种子制备同上,发酵时使用500 mL的三角弹簧瓶,每瓶装100 mL的发酵培养基,发酵总体积为5 L。于220 r/min、30 ℃培养7 d。发酵完成后,使用等体积的乙酸乙酯萃取3次,减压干燥,得到粗提物约10 g。
1.4 HPLC检测条件使用HPLC检测各菌株发酵产物次生代谢产物的差异情况,分析流动相为:乙腈-水(2‰的乙酸),流速:0.6 mL/min,均采用全波长扫描。HPLC洗脱条件:0-40 min:10%-50%乙腈梯度,40-50 min:50%-75%乙腈梯度,50-60 min:75%-100%乙腈梯度,60-68 min:纯乙腈洗脱,70-90 min:10%乙腈平衡柱子。色谱柱为:分析柱为Agilent 5 TC-C18(2) 250 mm×4.6 mm,SN:588925-902,PN:509550。进样体积为10 μL。
1.5 目标化合物的分离纯化利用10 g的60-100目层析硅胶将制备好的粗提物进行拌样。然后,使用60 g的200-300目层析硅胶进行干法装柱,层析柱的规格为:4 cm×60 cm。待上样完成后,使用三氯甲烷-甲醇为洗脱剂进行梯度洗脱。首先使用500 mL的氯仿进行洗脱,再逐渐增加洗脱剂中甲醇的比例,当洗脱剂中氯仿:甲醇的比例为20:1时(洗脱体积为1.5 L),利用HPLC检测该组分,发现了一个化合物与△PKS-3发酵样品中所缺少的化合物的保留时间和特征吸收波长均一致,该份样品命名为SF1。当洗脱剂氯仿:甲醇的比例为10:1时(洗脱体积为1.2 L),将洗脱下来的组分利用HPLC检测,发现含有△PKS-9中缺少的化合物,该份样品命名为SF2。
接下来,将含有目标化合物的两份样品,分别使用Sephadex LH-20进行凝胶过滤色谱纯化(色谱柱规格为1.8 cm×160 cm),洗脱剂为甲醇,流速为6-8 d/min,然后,分别将含有目标化合物的样品进行减压干燥后,继续分离纯化。其中,SF1组分利用正相硅胶柱层析(色谱柱规格为1.5 cm×50 cm),装柱硅胶为18.5 g,洗脱剂为石油醚:丙酮=85:15,分别收集洗脱组分,将含有较纯化合物的组分进行减压干燥,最终得到化合物SF1的纯品约100 mg。SF2组分再利用正相硅胶柱层析(色谱柱规格为1.5 cm×50 cm),装柱硅胶为15.0 g,利用氯仿:甲醇=10:1进行洗脱,将含有较纯的目标化合物组分进行减压干燥,最终得到化合物SF2的纯品为20 mg。
1.6 生物信息学预测antiSMASH在线预测网站预测基因组中次级代谢合成基因簇(http://antismash.secondarymetabolites.org/),通过frameplot 4.0 Beta预测基因簇的开放阅读框(http://biosyn.nih.go.jp/2ndfind/),美国国立生物技术信息中心NCBI网站上提供的BLAST软件(http://blast.ncbi.nlm.nih.gov/)进行蛋白预测。通过NRPS-PKS在线分析软件(http://www.nii.res.in/nrps-pks.html)对PKS各个模块及功能结构域进行预测[7]。
2 结果和分析2.1 野生型菌株与PKS中断突变株的次生代谢产物分析分别将野生型菌株S. albus DSM 41398和7个突变株(△PKS-1、△PKS-2、△PKS-3、△PKS-4、△PKS-5、△PKS-6和△PKS-9)在发酵培养基中培养7 d后取样,制备发酵粗提物样品。然后,利用HPLC全波长扫描,检测野生型和突变株发酵产物的差异性。当扫描波长为280 nm时,HPLC图谱表明野生型菌株和突变株△PKS-3的发酵产物在62 min处有一个明显的差异峰(图 1)。吸收波长特征分析显示,该化合物的最大吸收波长为226 nm,特征吸收峰的波长为276 nm。当检测波长为254 nm时,HPLC图谱表明突变株△PKS-9与野生型菌株的代谢物在53 min分钟左右有一个明显的差异峰,其最大吸收波长及特征吸收峰的波长均为254 nm (图 2)。本研究通过比较野生型菌株与突变菌株代谢产物的差异性,快速定位了2个目标差异峰,为下一步化合物的分离纯化奠定了基础。
|
| 图 1 野生型菌株和突变株次生代谢产物的HPLC分析(λ=280 nm)及目标峰的特征吸收谱Figure 1 HPLC analysis (λ=280 nm) of the secondary metabolites of wild type and mutant strains, together with the UV spectrum of the target compound. Wild type strain: S. albus DSM 41398; mutant strains: △PKS-1, △PKS-2, △PKS-3, △PKS-4, △PKS-5, △PKS-6 and △PKS-9. |
|
图选项
|
|
| 图 2 野生型菌株和突变株次生代谢产物的HPLC分析(λ=254 nm)及目标峰的特征吸收谱Figure 2 HPLC analysis (λ=254 nm) of the secondary metabolites of wild type and mutant strains, together with the UV spectrum of the target compound. Wild type strain: S. albus DSM 41398; mutant strains: △PKS-1, △PKS-2, △PKS-3, △PKS-4, △PKS-5, △PKS-6 and △PKS-9. |
|
图选项
|
2.2 野生型菌株的大量发酵与目标化合物的分离纯化
以上述2个HPLC图谱中的差异峰为导向,开展这2个化合物的分离纯化和结构鉴定研究。首先,对野生型菌株进行5 L的大量发酵,发酵完成后,利用乙酸乙酯萃取,制备粗提物样品。将粗样品用等量的60-100目硅胶进行拌样,然后再利用正相硅胶(200-300目)柱层析进行初步纯化,洗脱剂为氯仿-甲醇梯度洗脱,并采用HPLC分析寻找目标化合物,合并含有目标样品的洗脱液,进行减压干燥。接着,使用Sephadex LH-20分子筛层析纯化后,再利用正相硅胶柱层析进行交替纯化,直至得到目标化合物的纯品。最终得到化合物SF1的纯品约100 mg,化合物SF2的纯品约为20 mg。
2.3 目标化合物的结构鉴定放线吡喃酮的结构鉴定:纯化得到的化合物SF1为白色粉末,易溶于甲醇。称取5 mg纯品放入核磁管中,加入0.5 mL氘代甲醇进行溶解,利用Bruker Avance Ⅲ 600 MHz核磁共振仪收集核磁数据(表 1)。经1H-和13C-NMR的数据分析,该化合物的结构与放线吡喃酮(Actinopyranone)的相关数据较为一致。同时,该化合物的高分辨质谱(HR-ESI-MS)所得到的质荷比m/z为401.2732 ([M+H]+) (图 3),计算出的分子式为C25H37O4[8]。因此,将化合物SF1的结构鉴定为放线吡喃酮,结构式如图 4。
| No. | δH, muti. (J in Hz) | δC |
| 1 | 6.40 (d, 6.4) | 117.6 (d) |
| 2 | 6.49 (dd, 15.6/7.2) | 143.7 (d) |
| 3 | 2.45 (m) | 37.4 (d) |
| 4 | 2.15 (m) | 39.7 (t) |
| 5 | 5.48 (ddd, 15.0, 7.2/7.2) | 124.2 (d) |
| 6 | 6.03 (d, 15.2) | 137.1 (d) |
| 7 | / | 133.3 (s) |
| 8 | 5.21 (d, 9.0) | 134.5 (d) |
| 9 | 2.58 (m) | 36.2 (d) |
| 10 | 3.60 (d, 7.8) | 83.2 (d) |
| 11 | / | 136.7 (s) |
| 12 | 5.34 (m) | 121.3 (d) |
| 13 | 1.64 (s) | 11.7 (q) |
| 14 | 1.64 (s) | 9.8 (q) |
| 15 | 0.71 (d, 7.2) | 16.8 (q) |
| 16 | 1.72 (s) | 11.7 (q) |
| 17 | 1.05 (d, 6.6) | 18.5 (q) |
| 1′ | / | 153.0 (s) |
| 2′ | / | 116.8 (s) |
| 3′ | / | 181.8 (s) |
| 4′ | / | 98.6 (s) |
| 5′ | / | 162.8 (s) |
| 6′ | 1.88 (s) | 8.2 (q) |
| 7′ | 1.72 (s) | 5.7 (q) |
| 8′ | 4.00 (s) | 55.2 (q) |
|
| 图 3 放线吡喃酮的高分辨质谱检测结果Figure 3 The result of HR-ESI-MS analysis of actinopyranone. |
|
图选项
|
|
| 图 4 放线吡喃酮的化学结构Figure 4 The chemical structure of actinopyranone. |
|
图选项
|
洋橄榄菌素的结构鉴定:化合物SF2为白色粉末,易溶于氯仿。称取8 mg样品溶于氘代氯仿中,在核磁共振仪上进行核磁数据的收集(表 2)。高分辨质谱检测该化合物质荷比m/z为1047.5012 ([M+Na]+) (图 5),得到的分子式为C54H88O18Na。质谱数据和核磁共振数据与文献报道的基本相符[9]。通过1H-和13C-NMR的数据解析,将化合物SF2鉴定为洋橄榄菌素(Elaiophylin),结构如图 6。
| No. | δH, muti. (J in Hz) | δC |
| 1 | / | 167.1 (s) |
| 2 | 5.68 (1H, d, 15.6) | 121.3 (d) |
| 3 | 6.84 (1H, dd, 15.6/11.4) | 144.8 (d) |
| 4 | 6.11 (1H, dd, 15.0/11.4) | 130.6 (d) |
| 5 | 5.63 (1H, dd, 15.0/10.2) | 144.9 (d) |
| 6 | 2.51 (1H, m) | 41.2 (d) |
| 7 | 5.09 (1H, d, 10.8) | 75.8 (d) |
| 8 | 1.82 (1H, m) | 36.2 (d) |
| 9 | 3.81 (1H, m) | 68.3 (d) |
| 10 | 1.62 (1H, m) | 42.9 (d) |
| 11 | / | 99.2 (d) |
| 12 | 1.04 (1H, m) | 36.6 (t) |
| 13 | 2.28 (1H, m) | 69.4 (d) |
| 14 | 1.07 (1H, m) | 47.9 (d) |
| 15 | 3.79 (1H, m) | 66.4 (d) |
| 16 | 0.98 (3H, d, 6.6) | 18.8 (q) |
| 17 | 1.05 (3H, d, 6.0) | 15.5 (q) |
| 18 | 0.85 (3H, d, 7.2) | 9.6 (q) |
| 19 | 1.06 (3H, d, 6.6) | 7.0 (q) |
| 20 | 1.41 (1H, m) 1.62 (1H, m) |
19.2 (q) |
| 21 | 0.80 (3H, d, 6.0) | 8.8 (q) |
| 22 | 4.92 (1H, d, 3.6) | 92.5 (d) |
| 23 | 1.78 (2H, m) | 32.7 (t) |
| 24 | 3.81 (1H, m) | 64.9 (d) |
| 25 | 3.73 (1H, m) | 70.3 (d) |
| 26 | 4.27 (1H, m) | 65.8 (d) |
| 27 | 1.24 (3H, d, 6.8) | 17.1 (q) |
| 1′ | / | 167.1 (s) |
| 2′ | 5.68 (1H, d, 15.6) | 121.3 (d) |
| 3′ | 6.84 (1H, dd, 15.6/11.4) | 144.8 (d) |
| 4′ | 6.11 (1H, dd, 15.0/11.4) | 130.6 (d) |
| 5′ | 5.63 (1H, dd, 15.0/10.2) | 144.9 (d) |
| 6′ | 2.51 (1H, m) | 41.2 (d) |
| 7′ | 5.09 (1H, d, 10.8) | 75.8 (d) |
| 8′ | 1.82 (1H, m) | 36.2 (d) |
| 9′ | 3.81 (1H, m) | 68.3 (d) |
| 10′ | 1.62 (1H, m) | 42.9 (d) |
| 11′ | / | 99.2 (d) |
| 12′ | 1.04 (1H, m) | 36.6 (t) |
| 13′ | 2.28 (1H, m) | 69.4 (d) |
| 14′ | 1.07 (1H, m) | 47.9 (d) |
| 15′ | 3.79 (1H, m) | 66.4 (d) |
| 16′ | 0.98 (3H, d, 6.6) | 18.8 (q) |
| 17′ | 1.05 (3H, d, 6.0) | 15.5 (q) |
| 18′ | 0.85 (3H, d, 7.2) | 9.6 (q) |
| 19′ | 1.06 (3H, d, 6.6) | 7.0 (q) |
| 20′ | 1.41 (1H, m) 1.62 (1H, m) | 19.2 (q) |
| 21′ | 0.80 (3H, d, 6.0) | 8.8 (q) |
| 22′ | 4.92 (1H, d, 3.6) | 92.5 (d) |
| 23′ | 1.78 (2H, m) | 32.7 (t) |
| 24′ | 3.81 (1H, m) | 64.9 (d) |
| 25′ | 3.73 (1H, m) | 70.3 (d) |
| 26′ | 4.27 (1H, m) | 65.8 (d) |
| 27′ | 1.24 (3H, d, 6.8) | 17.1 (q) |
|
| 图 5 洋橄榄菌素的高分辨质谱检测结果Figure 5 The result of HR-ESI-MS analysis of elaiophylin. |
|
图选项
|
|
| 图 6 洋橄榄菌素的化学结构Figure 6 The chemical structure of elaiophylin. |
|
图选项
|
2.4 放线吡喃酮和洋橄榄菌素的生物合成途径分析
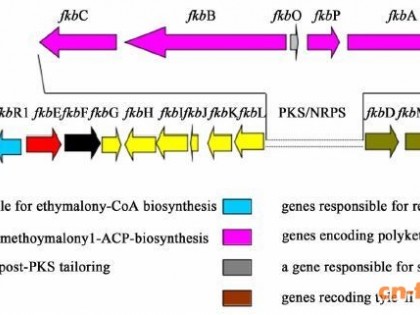
放线吡喃酮的生物合成途径分析:首先利用antiSMASH预测化合物的BGC,再使用frameplot 4.0 Beta预测基因簇每个开放阅读框,NCBI网站上的Blast软件进行蛋白的功能预测(表 3)。放线吡喃酮的BGC长度为104 kb,其生物合成的组装形式如图 7-A。在放线吡喃酮的BGC中含有5个聚酮合酶编码基因,编码了5个聚酮合酶蛋白,ATPN1含有起始模块和第1、2个延伸模块,AT识别的底物分别为丙二酰辅酶A、甲基丙二酰辅酶A、甲基丙二酰辅酶A;第1个延伸模块还含有KR、DH催化功能域,最终催化酮基变为C=C;第2个延伸模块含有KR催化功能域,最终催化酮基变为羟基。ATPN2含有第3、4个延伸模块,AT识别的底物分别为甲基丙二酰辅酶A、丙二酰辅酶A;第3、4个延伸模块都含有KR、DH催化功能域,最终催化2个酮基变为2个C=C键。ATPN3含有第5个延伸模块,AT识别的底物为甲基丙二酰辅酶A;第5个延伸模块还含有KR、DH、ER催化功能域,最终催化酮基变为C-C键。ATPN4含有第6、7个延伸模块,AT识别的底物分别为丙二酰辅酶A、甲基丙二酰辅酶A;第6个延伸模块还含有KR、DH催化功能域,最终催化酮基变为C=C键;第7个延伸模块还含有KR催化功能域,但其不发挥催化酮基变为羟基的功能。ATPN5含有第8个延伸模块,AT识别的底物为甲基丙二酰辅酶A;第8个延伸模块还含有KR催化功能域,但其不发挥催化酮基变为羟基的功能。除此之外还含有多个属于HxlR (orf1)、LuxR (orf2, orf36)或IclR (orf11)家族的调控基因以及ABC转运蛋白编码基因(orf6、orf7、orf8、orf9)。
| Gene | aa | Proposed function | Similar protein (acc. number) | Identity/% |
| orf1 | 147 | HxlR family transcriptional regulator | WP_040246795 | 99 |
| orf2 | 910 | LuxR family transcriptional regulator | WP_040406901 | 35 |
| orf3 | 273 | Trypsin | WP_009943017.1 | 44 |
| orf4 | 244 | Flavodoxin | WP_062990802.1 | 84 |
| orf5 | 276 | Amidohydrolase | WP_070198137.1 | 77 |
| orf6 | 337 | ABC transporter substrate-binding protein | WP_003935721.1 | 41 |
| orf7 | 270 | ABC transporter permease | WP_078564501.1 | 79 |
| orf8 | 403 | ABC transporter | WP_051431375.1 | 71 |
| orf9 | 215 | ABC transporter permease | WP_049570957.1 | 85 |
| orf10 | 310 | Alpha/beta hydrolase | WP_051307766.1 | 80 |
| orf11 | 255 | IclR family transcriptional regulator | WP_042189077 | 45 |
| orf12 | 275 | Hypothetical protein | WP_023530568.1 | 78 |
| orf13 | 495 | MFS transporter | WP_078617773.1 | 62 |
| orf14 | 120 | DNA translocase FtsK | WP_066735376.1 | 52 |
| atpn1 | 4575 | Polyketide synthase | WP_009946797.1 | 51 |
| atpn2 | 3746 | Type Ⅰ polyketide synthase | WP_052718869.1 | 52 |
| atpn3 | 2225 | Polyketide synthase | BAH02268.1 | 56 |
| atpn4 | 3438 | Type Ⅰ polyketide synthase | WP_040256246.1 | 57 |
| atpn5 | 1894 | Type Ⅰ polyketide synthase | WP_027745028.1 | 58 |
| orf15 | 206 | Threonylcarbamoyl-AMP synthase | WP_005321163.1 | 92 |
| orf16 | 695 | Regulator | 044366483 | 47 |
| orf17 | 171 | Hypothetical protein | WP_028430832.1 | 75 |
| orf18 | 144 | MFS transporter | WP_054228783.1 | 78 |
| orf19 | 300 | Transporter | WP_043498359 | 75 |
| orf20 | 477 | 3-carboxy-cis, cis-muconate cycloisomerase | WP_078076776.1 | 80 |
| orf21 | 660 | FAD-binding protein | WP_078076775.1 | 77 |
| orf22 | 421 | UDP-N-acetyl-D-glucosamine dehydrogenase | WP_078076774.1 | 88 |
| orf23 | 385 | Aminotransferase DegT | WP_078076773.1 | 80 |
| orf24 | 408 | Aminotransferase DegT | WP_069627421.1 | 84 |
| orf25 | 315 | 4-hydroxy-3-methylbut-2-enyl diphosphate reductase | WP_078079697.1 | 84 |
| orf26 | 355 | Polyprenyl synthetase | WP_078076771.1 | 78 |
| orf27 | 360 | Oxidoreductase | WP_033439990 | 45 |
| orf28 | 289 | Xylose isomerase | WP_031231149.1 | 81 |
| orf29 | 442 | Peptidase M20 | WP_079128120.1 | 78 |
| orf30 | 381 | Lactate dehydrogenase | WP_078076768.1 | 86 |
| orf31 | 305 | Oxidoreductase | WP_078076767.1 | 85 |
| orf32 | 160 | Hypothetical protein | WP_069627431.1 | 83 |
| orf33 | 607 | 1-deoxy-D-xylulose-5-phosphate synthase | WP_078076766.1 | 83 |
| orf34 | 384 | 4-hydroxy-3-methylbut-2-en-1-yl diphosphate synthase | WP_067371241.1 | 95 |
| orf35 | 279 | Streptogrisin A | SCE17834.1 | 90 |
| orf36 | 815 | LuxR family transcriptional regulator | AGK80163 | 38 |
|
| 图 7 放线吡喃酮生物合成基因簇(A)及生物合成途径推导(B)Figure 7 Biosynthetic gene cluster (A) and deduction of biosynthetic pathway (B) of actinopyranone. |
|
图选项
|
基于以上功能基因组成的分析,对放线吡喃酮的生物合成途径进行了推导(图 7-B)。从其生物合成途径可以看出1分子放线吡喃酮的合成需要1分子的乙酰辅酶A、2分子的丙二酰辅酶A和6分子的甲基丙二酰辅酶A,放线吡喃酮聚酮链合成成熟之后被硫酯酶(TE)从ACP上水解下来,经过环化和后修饰途径,最终形成放线吡喃酮。
洋橄榄菌素的生物合成途径分析:利用与放线吡喃酮BGC相同的分析策略,对洋橄榄菌素BGC中的相关功能蛋白进行分析,分析结果如表 4所示。其中,洋橄榄菌素的BGC全长为70 kb,功能基因的组装如图 8-A。洋橄榄菌素的BGC中含有5个聚酮合酶基因,编码5个聚酮合酶蛋白,ELPY5含有3个模块,包括了起始模块和第1、2个延伸模块,AT识别的底物分别为丙二酰辅酶A、乙基丙二酰辅酶A、丙二酰辅酶A;此外,第1个延伸模块还含有KR、DH催化功能域,但其DH催化结构域不发挥作用,最终催化酮基变为羟基;第2个延伸模块含有KR催化功能域,最终催化酮基变为羟基。ELPY4含有第3个延伸模块,AT识别的底物为甲基丙二酰辅酶A;第3个延伸模块还含有KR催化功能域,但其不发挥催化酮基变为羟基的作用。ELPY3含有第4个延伸模块,AT识别的底物为甲基丙二酰辅酶A;第4个延伸模块还含有KR催化功能域,最终催化酮基变为羟基。ELPY2含有第5、6个延伸模块,AT识别的底物分别为甲基丙二酰辅酶A、丙二酰辅酶A;第5个延伸模块还含有KR催化功能域,最终催化酮基变为羟基;第6个延伸模块还含有KR、DH催化功能域,最终催化酮基变为C=C键。ELPY1含有第7个延伸模块,AT识别的底物为丙二酰辅酶A;第7个延伸模块还含有KR、DH催化功能域,最终催化酮基变为C=C键。除此之外还含有2个属于LuxR家族的调控基因。由于洋橄榄菌素为二重内酯类结构,该类产物的释放机制较为特殊,首先是1个聚酮延伸完成的分子结合在TE结构域的活性位点上,然后,待另一分子的聚酮链组装完成后,TE结构域催化2个聚酮链发生分子间的亲和反应,形式二重内酯的结构[10]。
| Gene | aa | Proposed function | Similar protein (acc. number) | Identity/% |
| orf1 | 761 | Membrane protein | WP_029181812.1 | 56 |
| orf2 | 52 | Reguatory protein, luxR family | SCL73296.1 | 71 |
| orf3 | 486 | NDP-hexose 2, 3-dehydratase | AGP52909.1 | 63 |
| orf4 | 328 | Aldo/keto reductase | WP_078647113.1 | 79 |
| orf5 | 341 | NAD-dependent dehydratase | EXU62919.1 | 60 |
| orf6 | 216 | LuxR family transcriptional regulator | AGP52912.1 | 85 |
| orf7 | 458 | Signal transduction histidine kinase | SED09057.1 | 62 |
| orf8 | 245 | MULTISPECIES: ABC transporter | WP_014056964.1 | 84 |
| orf9 | 302 | Multidrug ABC transporter ATP-binding protein | WP_059148435.1 | 88 |
| orf10 | 209 | dTDP-4-dehydrorhamnose 3, 5-epimerase | WP_079259966.1 | 78 |
| orf11 | 417 | Glycosyltransferase | ADP68587.2 | 81 |
| orf12 | 261 | Oleoyl-ACP hydrolase | AGP52918.1 | 74 |
| elpy1 | 2086 | Polyketide synthase | WP_059148431.1 | 75 |
| elpy2 | 3407 | Polyketide synthase | AGP52920.1 | 76 |
| elpy3 | 1631 | Polyketide synthase | WP_079259961.1 | 77 |
| elpy4 | 1622 | Polyketide synthase | WP_078639065.1 | 71 |
| elpy5 | 4391 | Polyketide synthase | WP_009946797.1 | 53 |
| orf13 | 324 | dTDP-glucose 4, 6-dehydratase | WP_059145979.1 | 84 |
| orf14 | 306 | Glucose-1-phosphate thymidylyltransferase | AEM83455.1 | 85 |
| orf15 | 1125 | LuxR family transcriptional regulator | AQW47948.1 | 63 |
|
| 图 8 洋橄榄菌素生物合成基因簇(A)及生物合成途径推导(B)Figure 8 Biosynthetic gene cluster (A) and deduction of biosynthetic pathway (B) of elaiophylin. |
|
图选项
|
基于上述功能基因组成的分析,对洋橄榄菌素的生物合成途径进行了推导(图 8-B)。从其生物合成途径可以看出1分子洋橄榄菌素的合成需要2分子的乙酰辅酶A、6分子的丙二酰辅酶A、6分子的甲基丙二酰辅酶A和2分子的乙基丙二酰辅酶A,洋橄榄菌素聚酮链组装完成之后被硫酯酶(TE)从ACP上水解下来,经过环化和后修饰途径,最终形成洋橄榄菌素。
3 讨论和展望目前,以基因组序列导向的新结构微生物药物的发现技术已经得到了广泛的应用。此外,许多研究者还集成了基因组测序、生物信息学、分子生物学、合成生物学等生物学技术,发展了多样化的活性天然产物发掘策略[11-13],使新的微生物活性代谢产物的发现更加聚焦和便捷,也获得了很多的成功。一般说来,通过对目标基因簇的定向失活,使其失去合成目标产物的能力,将突变株和野生型的发酵代谢产物进行比较,从而找出目标化合物,进行结构鉴定和生物活性分析,依然是快速发现目标化合物的方法之一[5],本研究也利用该策略成功获得了具有生物活性的放线吡喃酮和洋橄榄菌素两个重要的聚酮类天然产物。
放线吡喃酮BGC的发现及生物合成途径的解析,将为本家族化合物生物合成的研究提供材料。同时,放线吡喃酮的分子结构与杀粉蝶菌素非常相似。最大的区别在于放线吡喃酮的聚酮链释放后,经过一步分子内环化,最终的分子中含有1个吡喃环。杀粉蝶菌素的聚酮链释放后,先是经过一步氨基的加载,然后才会发生分子内环化,形成含有1个吡啶环的结构。由于放线吡喃酮和杀粉蝶菌素均具有良好的生物活性[14],后续对于二者的比较研究或者分子结构的定向改造值得进一步探索和深入研究。
洋橄榄菌素具有很好的抗肿瘤活性[15],也受到了许多研究者的关注,具有很好的潜在应用价值,对于其产量的代谢工程优化和分子结构的遗传学改造均具有重要的现实意义。由于该菌株的相关操作已经非常成熟,所以利用该菌株来开展洋橄榄菌素的代谢工程优化也具有较强的可行性。同样,S. albus DSM 41398中含有多个Ⅰ型聚酮类化合物的BGCs,它们的表达与洋橄榄菌素的生物合成依然具有前体物的竞争情况。后续通过中断竞争性基因簇,使更多前体物流向洋橄榄菌素的生物合成,进而有可能进一步提高其产量,为药物研发提供物质保障。
由于盐霉素产生菌具有较强的天然产生聚酮类化合物的潜力,而且盐霉素的产量经过优化后已经达到了6.6 g/L[6]。本研究为后期对于PKS-1、PKS-2、PKS-4、PKS-5、PKS-6生物合成基因簇所对应化合物的发掘提供较好的研究示范。后续针对盐霉素及本研究中发现的放线吡喃酮和洋橄榄菌素的生物合成基因簇进行定向多重失活,减少底物的分流和菌株的负担,很可能会成为发现其他化合物更为有效的手段。
| [1] | Yao YP, Wang WS, Yang KQ. Efficient production of polyketide products in Streptomyces hosts-a review. Acta Microbiologica Sinica, 2016, 56(3): 418-428. (in Chinese) 姚永鹏, 王为善, 杨克迁. 链霉菌中高效生产聚酮化合物的研究方法及进展. 微生物学报, 2016, 56(3): 418-428. |
| [2] | Chen MJ, Wang GH, Dai SK, Xie LW, Li X. Polyketide antibiotics produced by polyketide synthase in Streptomyces-a review. Acta Microbiologica Sinica, 2009, 49(12): 1555-1563. (in Chinese) 陈敏捷, 王广华, 戴世鲲, 谢练武, 李翔. 链霉菌聚酮类次生代谢产物及其生物合成基因簇. 微生物学报, 2009, 49(12): 1555-1563. DOI:10.3321/j.issn:0001-6209.2009.12.002 |
| [3] | Doroghazi JR, Albright JC, Goering AW, Ju KS, Haines RR, Tchalukov KA, Labeda DP, Kelleher NL, Metcalf WW. A roadmap for natural product discovery based on large-scale genomics and metabolomics. Nature Chemical Biology, 2014, 10(11): 963-968. DOI:10.1038/nchembio.1659 |
| [4] | Jin C. Relationship of polyketide biosynthetic gene clusters. Microbiology China, 2014, 41(7): 1470. (in Chinese) 金城. 聚酮类抗生素生物合成基因簇间的关系. 微生物学通报, 2014, 41(7): 1470. |
| [5] | Zerikly M, Challis GL. Strategies for the discovery of new natural products by Genome Mining. Chem Bio Chem, 2009, 10(4): 625-633. DOI:10.1002/cbic.200800389 |
| [6] | Lu CY, Zhang XJ, Jiang M, Bai LQ. Enhanced salinomycin production by adjusting the supply of polyketide extender units in Streptomyces albus. metabolic Engineering, 2016, 35: 129-137.DOI:10.1016/j.ymben.2016.02.012 |
| [7] | Jiang CY, Wang HG, Kang QJ, Liu J, Bai LQ. Cloning and characterization of the polyether salinomycin biosynthesis gene cluster of Streptomyces albus XM211. Applied and Environmental Microbiology, 2012, 78(4): 994-1003. DOI:10.1128/AEM.06701-11 |
| [8] | Schleissner C, Pérez M, Losada A, Rodriguez P, Crespo C, Zúñiga P, Fernández R, Reyes F, de la Calle F. Antitumor actinopyranones produced by Streptomyces albus POR-04-15-053 isolated from a marine sediment. Journal of Natural Products, 2011, 74(7): 1590-1596. DOI:10.1021/np200196j |
| [9] | Zhang Y, Zhou X, Song YX, Chen YY, Huang HB, Li J, Hua Y, Ju JH. Elaiophylin from deep South China Sea-derived Streptomyces albiflaviniger SCSIO ZJ28. Natural Product Research and Development, 2013, 25(2): 185-189. (in Chinese) 张云, 周潇, 宋永相, 陈芸芸, 黄洪波, 李洁, 华燕, 鞠建华. 南海深海链霉菌Streptomyces albiflaviniger SCSIO ZJ28中Elaiophylin的分离鉴定. 天然产物研究与研发, 2013, 25(2): 185-189. |
| [10] | Zhou YJ, Prediger P, Dias LC, Murphy AC, Leadlay PF. Macrodiolide formation by the thioesterase of a modular polyketide synthase. Angewandte Chemie International Edition, 2015, 54(17): 5232-5235.DOI:10.1002/anie.v54.17 |
| [11] | Ziemert N, Alanjary M, Weber T. The evolution of genome mining in microbes-a review. Natural Product Reports, 2016, 33(8): 988-1005. DOI:10.1039/C6NP00025H |
| [12] | Thaker MN, Wang WL, Spanogiannopoulos P, Waglechner N, King AM, Medina R, Wright GD. Identifying producers of antibacterial compounds by screening for antibiotic resistance. Nature Biotechnology, 2013, 31(10): 922-927. DOI:10.1038/nbt.2685 |
| [13] | Jiang WJ, Zhu TF. Targeted isolation and cloning of 100-kb microbial genomic sequences by Cas9-assisted targeting of chromosome segments. Nature Protocols, 2016, 11(5): 960-975. DOI:10.1038/nprot.2016.055 |
| [14] | Liu Q, Yao F, Chooi YH, Kang QJ, Xu W, Li YR, Shao YC, Shi YF, Deng ZX, Tang Y, You DL. Elucidation of piericidin A1 biosynthetic locus revealed a thioesterase-dependent mechanism of α-pyridone ring formation. Chemistry & Biology, 2012, 19(2): 243-253. |
| [15] | Han Y, Tian EL, Xu DB, Ma M, Deng ZX, Hong K. Halichoblelide D, a new elaiophylin derivative with potent cytotoxic activity from mangrove-derived Streptomyces sp. 219807. Molecules, 2016, 21(8): 970. |