, 郑亚琴 , Yaqin Zheng
, 郑亚琴 , Yaqin Zheng 随着化石资源的不断消耗以及环境污染问题的加剧,利用微生物合成高附加值的工业产物受到愈来愈多的关注与重视[1]。在众多的平台化合物中,琥珀酸在食品、化工、农业等领域都具有非常广泛的应用[2-3]。琥珀酸还可以作为中间体合成γ-丁内酯、1, 4-丁二胺、四氢呋喃等化合物[4]。众多附加应用使得琥珀酸在2004年被美国能源部确定为最具价值的生物炼制产品[5]。
在微生物细胞内,琥珀酸是TCA循环的中间代谢产物,因此可利用生物质原料发酵法生产琥珀酸[6-7]。当前,利用大肠杆菌等基因工程菌生产琥珀酸可以达到较高的产率和产量,但是细菌不耐受酸及渗透压,需要添加碱液以维持中性的发酵环境[8-10]。发酵过程持续不断的pH值调节增加了染菌的概率。发酵结束后需要用酸调节pH值至酸性,使琥珀酸盐转变成琥珀酸,发酵成本大幅增加[11]。跟细菌等原核生物相比较,酵母能耐受低pH值,增加了其在工业应用的优势。
解脂耶氏酵母Yarrowia lipolytica是非传统酵母,严格好氧,利用完整的TCA循环和电子传递链维持生长[12-13]。它能积累大量的有机酸,包括柠檬酸、异柠檬酸、α-酮戊二酸等[14-16]。经过代谢工程改造后,敲除琥珀酸脱氢酶(Succinate dehydrogenase,SDH)的编码基因或者弱化该基因的启动子,Y. lipolytica可积累琥珀酸[17-18]。2010年Gao等通过敲除Sdh2亚基成功构建可积累4.0 g/L琥珀酸的Y. lipolytica重组菌,随后采用化学诱变的方法获得琥珀酸产量进一步提高的突变株。我们课题组通过基因敲除Y. lipolytica Po1f的SDH的Sdh5亚基获得了重组菌PGC01003[19]。PGC01003在不控制pH值的中性环境下以甘油作碳源发酵足够长时间能生产琥珀酸,这充分证明了该菌株的鲁棒性。然而,实验存在一些亟待解决的问题。重组菌在发酵产生琥珀酸的同时,由于代谢溢出,还产生较多的副产物乙酸。乙酸对菌体具有毒害作用,抑制细胞生长和琥珀酸发酵。如果不控制发酵液的pH值,琥珀酸的产量很低[19]。
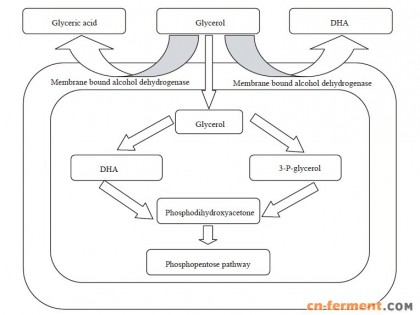
为实现低pH值发酵生产琥珀酸,我们对Y. lipolytica乙酸代谢的可能途径进行分析,并通过代谢工程的策略降低乙酸的溢出(图 1)。首先干扰旁路代谢(Pyruvate dehydrogenase bypass,PDH bypass),基因敲除丙酮酸脱羧酶(Pyruvate decarboxylase,PDC)基因或者过量表达乙酰辅酶A合酶(Acetyl coenzyme A synthetase,ACS),分析重组菌的乙酸产量变化。随后基因敲除乙酰辅酶A水解酶(Acetyl-CoA hydrolase,ACH1),获得重组菌PGC11505,乙酸分泌量大幅降低。
|
| 图 1 解脂酵母Y. lipolytica的主要中心代谢途径Figure 1 Central pathways related to succinic acid production of Y. lipolytica. |
|
图选项
|
解脂酵母Y. lipolytica Po1f (URA3-,Leu2-)由法国INRA实验室的Madzak Catherine教授馈赠。产琥珀酸的重组酵母菌株均为作者所在实验室构建和保存[20],详见表 1。
| Strain name | Description |
| PGC01003 | Y. lipolytica Po1f derivate, sdh5 |
| PGC11201 | Y. lipolytica Po1f derivate, sdh5, pdc |
| PGC11301 | Y. lipolytica Po1f derivate, sdh5, Ylacs |
| PGC11401 | Y. lipolytica Po1f derivate, sdh5, oSeacs |
| PGC11505 | Y. lipolytica Po1f derivate, sdh5, ach1 |
酵母种子培养和发酵培养基为YPG完全培养基,含20 g/L甘油,20 g/L蛋白胨,10 g/L酵母粉,自然pH。制备固体培养基时添加2%的琼脂。
1.3 培养方法1.3.1 种子培养挑取生长良好的单菌落接种于含50 mL的YPG液体培养基的250 mL三角瓶中,28 ℃、220 r/min培养24 h。
1.3.2 乙酸耐受培养将种子培养液按1%接种量转接至含不同浓度乙酸(1、2、5 g/L)的YPG液体培养基中,28 ℃、220 r/min培养24-48 h。
1.3.3 摇瓶发酵培养将种子培养液按1%接种量转接至50 mL的发酵培养基YPG中,28 ℃、220 r/min培养,定时取样测定菌体OD600、甘油和有机酸的含量。
1.4 分析检测菌体浓度采用分光光度计测600 nm下的吸光度。采用HPLC定量检测发酵液中的甘油和有机酸的含量。HPLC分析条件:流动相为5 mmol/L稀硫酸,色谱柱为Aminex HPX-87H (Bio-Rad,USA),流速0.6 mL/min,柱温65 ℃,进样量10 μL,检测器为示差折光检测器。
2 结果与分析2.1 重组菌PGC01003的琥珀酸发酵前期,基因敲除了解脂酵母Po1f菌株中参与TCA循环的琥珀酸脱氢酶(Succinate dehydrogenase,SDH)的Sdh5亚基编码基因Ylsdh5,成功构建了琥珀酸脱氢酶失活的突变株,获得可积累琥珀酸的解脂酵母重组菌PGC01003[19]。图 2分别列出了重组菌PGC01003和对照菌Po1f在YPG摇瓶发酵时甘油浓度、菌体OD值、琥珀酸、乙酸、柠檬酸的产量。可以看到,重组菌PGC01003从24 h开始菌体生长减缓,30 h达到稳定期,OD值最大为14.0,发酵结束时甘油仍残余3.8 g/L。而Po1f可以持续生长至72 h,直至甘油消耗完全方停止生长,OD值达到33.5。Po1f的发酵产物主要是柠檬酸(5.0 g/L),只有极少量的琥珀酸和乙酸产生,PGC01003几乎不分泌柠檬酸(0.4 g/L),产生5.0 g/L琥珀酸。由于代谢流量不平衡,PGC01003发生代谢溢出,产生大量的副产物乙酸,在24 h时生成2.8 g/L乙酸,30 h时继续增加至4.8 g/L,48 h时乙酸浓度高达6.1 g/L,发酵终止。
|
| 图 2 解脂酵母PGC01003 (A)和Po1f (B)的琥珀酸发酵比较Figure 2 Fermentation profiles of PGC01003 (A) and Po1f (B) in YPG flasks. Squares stand for OD600, circles stand for glycerol concentrations, diomands stand for succinic acid concentrations, upright triangles stand for acetic acid concentrations and inverted triangles stand for citric acid concentrations. |
|
图选项
|
为检验增加溶氧是否有助于改善重组菌PGC01003的生长和琥珀酸生成,我们采用不带/带挡板的摇瓶以及不同装液量进行发酵(图 3)。由图 3A可以看出,由于溶氧的增加,使用带挡板的摇瓶可以提高对照菌Po1f的菌体生长,其OD值由34.0增至71.0,增长了1倍。然而,装液量为30 mL和50 mL时是否带挡板对PGC01003的生物量影响较少;装液量提高到80 mL以上时,PGC01003在带挡板的摇瓶中生长较好,可获得更高的生物量。此外,在装液量相同的发酵液中带挡板摇瓶的琥珀酸产量略有降低(图 3B),这可能是增加的溶氧促进了菌体的前期生长和维持所致。通过改变装液量或采用带挡板的摇瓶来改变发酵液溶氧的方式不能提高重组菌的琥珀酸产量。
|
| 图 3 PGC01003在不带/带挡板的摇瓶以及不同装液量条件下菌体OD值(A)和琥珀酸产量分析(B)Figure 3 OD (A) and succinic acid fermentation (B) of PGC01003 in flasks without/with baffles and with varied volumes of fermentation broth. |
|
图选项
|
为提高琥珀酸的产量,首先采用代谢工程干扰PDH旁路的策略阻断乙酸的溢出。PDH旁路由丙酮酸脱羧酶催化细胞质丙酮酸脱羧生成乙醛,乙醛在乙醛脱氢酶的作用下氧化为乙酸。细胞质的乙酸可以进一步被乙酰辅酶A合酶(Acetyl-CoA synthase,ACS)转化为乙酰辅酶A。乙酰辅酶A作为细胞内的高能代谢物,参与许多重要的合成和分解代谢[21]。本实验分别采用基因敲除丙酮酸脱羧酶编码基因Ylpdc、过量表达ACS的方法降低乙酸的浓度。
敲除PGC01003的Ylpdc基因获得重组菌株PGC11203,PGC11203在YPG培养基发酵72 h仍产生较高的乙酸(表 2),琥珀酸的产量和菌体OD略有下降,Ylpdc基因的缺失不但没有降低乙酸的溢出,而且还影响菌体的生长和琥珀酸发酵。另一替代策略是在PGC01003中过量表达ACS,将细胞质中多余的乙酸转化为高能化合物乙酰辅酶A。分别表达了Y. lipolytica自身的ACS和来自沙门氏菌Salmonella enterica的ACS,所构建的新菌株分别是PGC11301 (Ylacs)与PGC11401 (oSEacsL641P)。文献报道,ACS的活性受到翻译后乙酰化与去乙酰化系统修饰。而将S. enterica的ACS蛋白质中第641位的脯氨酸替换为亮氨酸可阻止其乙酰化,进而维持其活性状态[21]。根据Y. lipolytica的密码子偏好优化了SEacsL641P,并合成全新的基因序列oSeacs用于在Y. lipolytica的表达,其发酵情况如表 2所示。PGC11301的发酵情形与PGC01003相当,表明直接表达Y. lipolytica自身的ACS不能有效控制乙酸的溢出。相比之下,异源表达oSEacsL641P的重组菌PGC11401的菌体OD600增加至14.4,琥珀酸产量为6.3 g/L (提高了14.5%),乙酸产量降至4.6 g/L,下降了24.6%。对PDH旁路途径的扰动策略中,仅表达异源的oSEacsL641P降低了乙酸溢出,进而改善菌体生长,增加琥珀酸生产。经过点突变的oSEacsL641P能有效保持活性状态,催化乙酸生成乙酰辅酶A。
| Strains | Glycerol (g/L) | OD600 | Acetic acid titer (g/L) | Succinic aicd titer (g/L) | Succinic aicd yield (g/g) |
| PGC01003 | 22.0 | 13.95 | 6.1 | 5.5 | 0.25 |
| PGC11203 | 20.4 | 12.25 | 5.9 | 4.7 | 0.23 |
| PGC11301 | 20.9 | 12.30 | 5.7 | 4.8 | 0.23 |
| PGC11401 | 23.9 | 14.44 | 4.6 | 6.9 | 0.28 |
异源表达oSEacsL641P降低了乙酸产出,但是重组菌PGC11401仍溢出较多的乙酸,造成碳源的浪费,并影响细胞的正常生长。因此,我们推测Y. lipolytica还存在其他的乙酸溢出路径。经过代谢途径挖掘和文献分析发现酿酒酵母Saccharomyces cerevisiae的乙酰辅酶A水解酶(Acetyl-CoA hydrolase,ACH1)能催化乙酰辅酶A为乙酸和辅酶A[22]。随后,研究者发现该酶位于线粒体中,它具备辅酶A转移酶活性,可将乙酰辅酶A的乙酰基转移至琥珀酸生成乙酸和琥珀酰辅酶A[23]。通过基因序列比对与查找,我们获得Y. lipolytica的乙酰辅酶A水解酶编码基因Ylach1 (YALI0E30965g),基因敲除Ylach1构建了重组菌PGC11505。图 4A显示,发酵96 h重组菌PGC11505的琥珀酸产量达到7.0 g/L,比PGC01003提高了27.3%,乙酸只有0.48 g/L,菌体的生长接近对照菌Po1f (OD600达到29.4)。甘油被完全消耗掉,琥珀酸的转化率达到0.30 g/g。ACH1的失活阻断了线粒体中乙酰辅酶A转化为乙酸,同时也降低了琥珀酸向琥珀酰辅酶A的转化。乙酸溢出大幅度减少,这有利于菌体生长,提高了琥珀酸的产量。
|
| 图 4 PGC11505琥珀酸发酵分析(A)与乙酸耐受实验(B)Figure 4 Succinic acid fermentation profiles of PGC11505 (A) and effect of exogenous acetic acid on cell growth of PGC11505 (B). (A) Squares stand for OD600; circles stand for glycerol concentrations; diamonds stand for succinic acid concentrations and upright triangles stand for acetic acid concentrations. |
|
图选项
|
为进一步验证乙酸对Y. lipolytica细胞生长的影响,向培养基中外源添加乙酸,检测重组菌PGC11505的菌体生长。图 4B显示乙酸浓度为1 g/L即对PGC11505产生抑制作用,菌体的OD值在24 h和48 h分别降低了64.9%和66.0%。随着乙酸浓度的增加,细胞生长抑制愈加明显。当外源添加乙酸浓度为5 g/L时,细胞的生长被完全抑制。
3 结语琥珀酸作为一种高附加值有机酸,具有重要的生理功能。目前,细菌生产琥珀酸可以达到较高的产率和产量,但是细菌不耐受酸及渗透压,发酵过程需要添加碱液以维持中性环境,发酵结束后则需要用酸调节pH值至酸性,使琥珀酸盐转变成琥珀酸。酵母能耐受低pH值和环境压力,增加了其在工业应用的优势。解脂酵母是非传统酵母,能积累柠檬酸等大量有机酸。由于代谢不平衡,通过失活琥珀酸脱氢酶所获得的重组解脂酵母产生大量副产物乙酸[18-19]。乙酸的溢出对酵母细胞产生毒害作用,抑制菌体生长和发酵。通过干扰旁路途径,异源表达来自鼠沙门氏菌的乙酰辅酶A合酶可降低乙酸的溢出,乙酸产量降至4.6 g/L,下降了24.6%。而基因敲除乙酰辅酶A水解酶编码基因,乙酸的溢出降低至0.4 g/L,琥珀酸产量提高到7.0 g/L。乙酸溢出的有效控制恢复了菌体生长能力,为进一步的代谢工程构建高产琥珀酸的细胞工厂奠定基础。
| [1] | Liang QF, Qi QS. From a co-production design to an integrated single-cell biorefinery. Biotechnol Adv, 2014, 32(7): 1328–1335. DOI: 10.1016/j.biotechadv.2014.08.004 |
| [2] | Zheng P, Dong JJ, Sun ZH, et al. Fermentative production of succinic acid from straw hydrolysate by Actinobacillus succinogenes. Bioresour Technol, 2009, 100(8): 2425–2429. DOI: 10.1016/j.biortech.2008.11.043 |
| [3] | Nattrass L, Aylott M, Higson A. NNFCC renewable chemicals factsheet: Succinic acid. NNFCC 2013. |
| [4] | Mckinlay JB, Vieille C, Zeikus JG. Prospects for a bio-based succinate industry. Appl Microbiol Biotechnol, 2007, 76(4): 727–740. DOI: 10.1007/s00253-007-1057-y |
| [5] | Werpy T, Petersen G, Aden A, et al. Top value added chemicals from biomass, volume 1: results of screening for potential candidates from sugars and synthesis gas. Washington, DC: US Department of Energy, 2004. |
| [6] | Vuoristo KS, Mars AE, Sanders JP, et al. metabolic engineering of TCA cycle for production of chemicals. Trends Biotechnol, 2016, 34(3): 191–197. DOI: 10.1016/j.tibtech.2015.11.002 |
| [7] | Yin X, Madzak C, Du G, et al. Enhanced alpha-ketoglutaric acid production in Yarrowia lipolytica WSH-Z06 by regulation of the pyruvate carboxylation pathway. Appl Microbiol Biotechnol, 2012, 96(6): 1527–1537. DOI: 10.1007/s00253-012-4192-z |
| [8] | Borges ER, Pereira N. Succinate production from sugarcane bagasse hemicellulose hydrolysate by Actinobacillus succinogenes. J Ind Microbiol Biot, 2010, 38(8): 1001–1011. |
| [9] | Li YK, Li MJ, Zhang X, et al. A novel whole-phase succinate fermentation strategy with high volumetric productivity in engineered Escherichia coli. Bioresource Technol, 2013, 149: 333–340. DOI: 10.1016/j.biortech.2013.09.077 |
| [10] | Wang C, Cai H, Chen Z, et al. Engineering a glycerol utilization pathway in Corynebacterium glutamicum for succinate production under O2 deprivation. Biotechnol Lett, 2016, 38(10): 1791–1797. DOI: 10.1007/s10529-016-2166-4 |
| [11] | Jansen ML, van Gulik WM. Towards large scale fermentative production of succinic acid. Curr Opin Biotechnol, 2014, 30: 190–197. DOI: 10.1016/j.copbio.2014.07.003 |
| [12] | Nicaud JM. Yarrowia lipolytica. Yeast, 2012, 29: 409–418. DOI: 10.1002/yea.v29.10 |
| [13] | Liu HH, Ji XJ, Huang H. Biotechnological applications of Yarrowia lipolytica: past, present and future. Biotechnol Adv, 2015, 33(8): 1522–1546. DOI: 10.1016/j.biotechadv.2015.07.010 |
| [14] | Zhu Q, Jackson EN. metabolic engineering of Yarrowia lipolytica for industrial applications. Curr Opin Biotechnol, 2015, 36: 65–72. DOI: 10.1016/j.copbio.2015.08.010 |
| [15] | Gon alves FA, Colen G, Takahashi JA. Yarrowia lipolytica and its multiple applications in the biotechnological industry. Scientific World J, 2014, 1: 476207. |
| [16] | RywinʼSka A, Juszczyk P, Wojtatowicz M, et al. Glycerol as a promising substrate for Yarrowia lipolyticabiotechnological applications. Biomass Bioenerg, 2013, 48(1): 148–166. |
| [17] | Jost B, Holz M, Aurich A, et al. The influence of oxygen limitation for the production of succinic acid with recombinant strains of Yarrowia lipolytica. Appl Microbiol Biotechnol, 2015, 99(4): 1675–1686. DOI: 10.1007/s00253-014-6252-z |
| [18] | Yuzbashev TV, Yuzbasheva EY, Sobolevskaya TI, et al. Production of succinic acid at low pH by a recombinant strain of the aerobic yeast Yarrowia lipolytica. Biotechnol Bioeng, 2010, 107(4): 673–682. DOI: 10.1002/bit.22859 |
| [19] | Gao CJ, Yang XF, Wang HM, et al. Robust succinic acid production from crude glycerol using engineered Yarrowia lipolytica. Biotechnol Biofuels, 2016, 9(1): 179–189. DOI: 10.1186/s13068-016-0597-8 |
| [20] | Cui ZY, Gao CJ, Li JJ, et al. Engineering of unconventional yeast Yarrowia lipolytica for efficient succinic acid production from glycerol at low pH. metab Eng, 2017, 42: 126–133. DOI: 10.1016/j.ymben.2017.06.007 |
| [21] | Starai VJ, Gardner JG, Escalantesemerena JC. Residue Leu-641 of Acetyl-CoA synthetase is critical for the acetylation of residue Lys-609 by the Protein acetyltransferase enzyme of Salmonella enterica. J Biol Chem, 2005, 280(28): 26200–26205. DOI: 10.1074/jbc.M504863200 |
| [22] | Buu LM, Chen YC, Lee FJ. Functional characterization and localization of acetyl-CoA hydrolase, Ach1p, in Saccharomyces cerevisiae. J Biol Chem, 2003, 278(19): 17203–17209. DOI: 10.1074/jbc.M213268200 |
| [23] | Chen Y, Zhang Y, Siewers V, et al. Ach1 is involved in shuttling mitochondrial acetyl units for cytosolic C2 provision in Saccharomyces cerevisiae lacking pyruvate decarboxylase. FEMS Yeast Res, 2015, 15(3): 1–8. |