


 13720223167
13720223167



摘要:莽草酸是莽草酸代谢途径的中间产物,是合成手型药物的关键原料,也是合成芳香族类氨基酸(L-苯丙氨酸、L-酪氨酸、L-色氨酸)、泛醌、叶酸、维生素K2等芳香族化合物的关键中间体。通过基因改造微生物莽草酸代谢途径,可以使其大量积累莽草酸,进而为生物体内合成芳香族氨基酸奠定基础。本文综述了莽草酸代谢途径在重组大肠杆菌中的生物合成途径的研究进展,分析了莽草酸合成过程中关键酶基因改造对莽草酸积累的影响、辅因子再生对目的产物的影响,还讨论了减少莽草酸工程菌代谢副产物产生量的措施。随着代谢工程技术和合成生物学技术的发展,通过基因技术定向改造代谢途径,构建理想的高产菌株已经成为研究的新热点。
关键词:莽草酸代谢途径;基因改造;关键酶基因;辅因子
中图分类号: Q945.1 文献标志码: A 文章编号:1002-1302(2019)07-0019-05
莽草酸及其衍生物具有抗病毒、抗肿瘤、抗菌抗炎、抗血栓及抗脑缺血等多种药理活性。近年来,莽草酸作为临床上对抗禽流感的唯一有效药物达菲(主要成分为磷酸奥司他韦)的合成前体而倍受关注[1]。莽草酸途径广泛存在于植物、细菌等微生物中[2-5]。目前,莽草酸的工业化生产方式主要有2种:(1)从八角茴香的果实中提取[6];(2)通过化学合成的方式人工合成莽草酸[7]。从八角茴香中提取莽草酸,由于受到植物生长季节、生长周期、地域环境的限制,从而严重制约了莽草酸的大规模工业化生产;化学合成法由于生产工艺复杂、成本高、纯度不高等因素,也严重限制了莽草酸的规模化生产。利用微生物工程菌合成莽草酸,在生产规模、生产工艺与生产周期等方面具有显著优势,因此成为国内外很多医药公司与实验室研究的热点。目前已报道的莽草酸基因工程菌主要有大肠杆菌(Escherichia coli)[8]、枯草芽孢杆菌(Bacillus subtilis)[9]、谷氨酸棒状杆菌(Corynebacterium glutamicum)[10-11]、弗氏柠檬酸杆菌(Citrobacter freundii)[12-15]以及巨大芽孢杆菌(Bacillus megaterium)[16]等。
1 微生物生产莽草酸的研究进展
目前,利用微生物生物转化合成莽草酸的研究进展迅速,相关报道显示,目前工程菌株产莽草酸的最高产量达到 84 g/L。近几年来研究者构建的莽草酸工程菌株及其功能、产量等信息见表1。

2 莽草酸途径概述
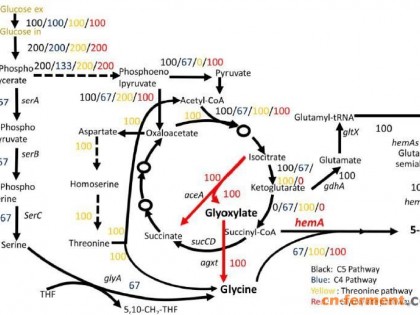
莽草酸途径(shikimic acid pathway)是以磷酸烯醇式丙酮酸、赤藓糖-4-磷酸为起始底物,在生物体内经过一系列催化酶的催化作用生成莽草酸,然后再以莽草酸为中间化合物合成一系列芳香族类化合物,如酪氨酸、苯丙氨酸、色氨酸等的过程。生物细胞以葡萄糖为初始底物,在經过糖酵解途径合成磷酸烯醇式丙酮酸的同时,通过戊糖磷酸途径合成赤藓糖-4-磷酸。生物体内的莽草酸途径以磷酸烯醇式丙酮酸与赤藓 糖-4-磷酸为起始化合物,在3-脱氧-阿拉伯庚酮糖-7- 磷酸(3-deoxy-D-arabinoheptulosonate-7-phosphate,简称DAHP)合成酶的作用下缩合形成3-脱氧-阿拉伯庚酮糖-7-磷酸。DAHP经过3-脱氢奎尼酸(3-dehydroquinic acid,简称DHQ)合酶的催化作用生成3-脱氢奎尼酸,DHQ在3-脱氢奎尼酸脱水酶的作用下生成3-脱氢莽草酸(3-dehydroshikimic acid,简称DHS)。DHS在莽草酸脱氢酶的作用下生成莽草酸(shikimic acid,简称SA)。莽草酸经莽草酸激酶催化生成莽草酸-3-磷酸(shikimate-3-phosphate,简称S3P)。随后,在一系列酶的催化下,经过分支酸(chorismic acid,简称CHA)分别转化为芳香族氨基酸苯丙氨酸、酪氨酸、色氨酸[24]。微生物细胞内的莽草酸代谢途径如图1所示[25]。
3 关键酶基因的过表达
通过质粒强化表达莽草酸代谢途径的限速酶基因,可以增强莽草酸代谢途径中的碳代谢流[17]。大肠杆菌莽草酸代谢途径中的3-脱氧-D-阿拉伯庚酮糖-7-磷酸(3-deoxy-D-arabino-heptulosonate-7-phosphate,简称DAHP)合酶(aroG、aroF、aroH为DAHP的3个同功异构酶)、3-脱氢奎尼酸合成酶(aroB)和莽草酸脱氢酶(aroE)是莽草酸代谢途径中的关键酶。DAHP合酶是由aroG、aroF和aroH基因编码的同工酶,分别受到芳香族类氨基酸L-苯丙氨酸、L-酪氨酸和L-色氨酸的反馈抑制。通过定点突变、随机诱变与突变株筛选等方法可以获取抗反馈抑制的3-脱氧-D-阿拉伯庚酮糖-7-磷酸合酶。为了解除终产物酪氨酸对aroG基因的反馈抑制,需要将其第146位氨基酸残基天冬氨酸替换为天冬酰胺,能够解除酪氨酸对aroG基因的反馈抑制,用亮氨酸残基替换DAHP合酶同工酶aroF的第148位氨基酸残基脯氨酸时,得到的突变菌株不受终产物苯丙氨酸的反馈抑制。因此,可以通过质粒表达或基因重组等方式过表达对酪氨酸反馈抑制不敏感的aroGfbr基因与对苯丙氨酸不敏感的aroFfbr基因,提高莽草酸代谢途径的碳代谢流。Draths等在莽草酸激酶完全缺失的大肠杆菌体内过表达aroFfbr基因,同时通过质粒过表达3-脱氢奎尼酸合成酶与莽草酸合成酶基因补料分批发酵培养42 h,使得莽草酸的产量达到27.2 g/L[26]。将这3个关键酶基因通过多个质粒或者1个共表达载体进行过表达,已经成为一套成熟的莽草酸工程菌改造策略。编码3-脱氢奎尼酸合成酶的基因aroB与抗反馈抑制的3-脱氧-D-阿拉伯庚酮糖-7-磷酸合成酶基因共表达,可以增强3-脱氢奎尼酸合成酶的活性,减少DAHP的过度积累,增强莽草酸代谢途径的碳代谢流。
4 中心代谢途径的改造
在微生物体内,葡萄糖经过糖酵解途径合成磷酸烯醇式丙酮酸(PEP)和赤藓糖-4-磷酸(E4P),PEP与E4P构成莽草酸代谢途径的起始物。因此,通过对中心代谢途径进行遗传改造,加强前体PEP与E4P的供应量,可以强化莽草酸代谢途径的碳流量。在非磷酸烯醇式丙酮酸:碳水化合物磷酸转移酶系统(简称PTS系统)的半乳糖透性酶(GalP)、半乳糖激酶(Galk)协同作用途径中以腺嘌呤核苷三磷酸(ATP)作为磷酸基团供体,不消耗PEP[8]。有研究表明,在 PTS+ 与PTS-Glc+中进入EMP和PPP途径的碳代谢流分别为766%、223%和93.1%、5.3%,表明PTS-galP+glk+ 能够促进EMP(糖酵解途径)中PEP的合成,提高莽草酸代谢途径中的碳流量。PTS系统中的EIIAGlc拥有碳代谢阻遏效应(carbon catabolic repression,简称CCR)的活性中心,可以阻遏细胞对葡萄糖以外其他碳源的利用分解,PTS-系统能够遏制CCR,使细胞能够充分利用多种碳源,强化莽草酸代谢途径的碳流量。
在细菌内,葡萄糖也可通过半乳糖透性酶介导的活性转运系统转运至细胞内,同时需要葡萄糖激酶蛋白Glk(由glk编码)对细胞内的葡萄糖进行磷酸化。因此,可以在PTS转运系统缺陷型菌株内加强表达GalP、Glk蛋白,以实现葡萄糖的转运功能。Yi等在PTS转运系统缺失的大肠杆菌体内通过质粒表达来自运动发酵单胞菌(Zymomonas mobilis)的葡萄糖易化体蛋白(glucose facilitator,简称Glf),结果显示,表达Glk、Glf的PTS缺陷型菌株经过48 h发酵培养,3-脱氢莽草酸的产量为60 g/L,与此同时,表达GalP、Glk的PTS缺陷型菌株经过60 h发酵培养,3-脱氢莽草酸的产量也为60 g/L,说明Glf系统也是一种比较有效的葡萄糖转运系统[27]。Chandran等在PTS转运系统缺陷型宿主菌体内通过质粒表达来自Zymomonas mobilis的葡萄糖易化体蛋白Glf与葡萄糖激酶蛋白Glk,同时解除DAHP合成酶的反馈抑制调节,过表达关键酶基因磷酸烯醇式丙酮酸合成酶基因(phosphoenolpyruvate synthase,简称ppsA),结果显示,在添加10 L酵母提取物的发酵罐中,莽草酸的产量达到了87 g/L,莽草酸的得率为33%[21]。
4.1 PTS系统的改造
磷酸烯醇式丙酮酸磷酸转移酶系统是大肠杆菌在细胞内参与葡萄糖从膜间质转运和磷酸化到细胞内的主要活性转运系统。在以葡萄糖为碳源的培养基内,PTS系统消耗了大约50%的磷酸烯醇式丙酮酸,而磷酸烯醇式丙酮酸是莽草酸代谢途径的起始物质,因此,合理改造PTS系统有利于提高莽草酸代谢途径前体PEP的利用率,从而提高莽草酸代谢途径的碳流量。Chandran等将运动发酵单胞菌来源的、以ATP作为磷酸基团供体的葡萄糖转运系统以质粒方式转化到PTS系统缺失的大腸杆菌内,同时过表达tktA基因以补料分批发酵培养,结果表明,莽草酸的产率达到 0.33 mol/mol(以单位量的葡萄糖产莽草酸的量计,下同),产量为84 g/L;同时,对于未进行PTS改造的菌株,同样采用补料分批发酵培养的莽草酸的积累量为62 g/L[21]。
4.2 增加莽草酸途径起始底物E4P的浓度
大量研究表明,过量表达DAHP合成酶并不能无限制地提高莽草酸代谢途径中的碳代谢流,必须同时加强表达tktA基因编码的转酮醇酶A,以增加磷酸戊糖途径中赤藓糖-4-磷酸的供应量,进而增加莽草酸代谢途径中的碳代谢流。Knop等在大肠杆菌体内过表达tktA基因后,莽草酸产量由 38 g/L 提高到52 g/L,莽草酸的产率由0.12 mol/mol上升到0.18 mol/mol[6]。有研究者在aroL、aroK双基因敲除菌株W3110(ΔaroLΔaroK)中转入含有莽草酸代谢途径中关键基因aroF、aroE、aroB、ppsA以及tktA基因的重组表达质粒pSUFEBPT,用补料分批发酵的方式发酵培养128 h,结果显示,莽草酸的产量达到39.3 g/L。
5 降低副产物的产量
莽草酸代谢途径中存在多步分支途径与可逆反应,代谢过程中产生的副产物奎尼酸(quinic acid,简称QA)和3-脱氢莽草酸对于细胞的生长有强烈的抑制作用,同时能够竞争性消耗碳源,从而严重降低莽草酸的产率与纯度。Knop等构建的大肠杆菌菌株SP1.1/pKD12.112发酵42 h,能够产生27.2 g/L莽草酸,但是副产物奎尼酸、3-脱氢莽草酸的产量分别有12.6、4.4 g/L,副产物的积累减少了莽草酸的含量与纯度,从而影响下游代谢反应[6]。YdiB是一种双功能酶,具有脱氢奎尼酸脱氢酶(aroD基因产物)和莽草酸脱氢酶(aroE基因产物)的双重功能,ydiB基因缺失后,奎尼酸合成受到限制,3-脱氢奎尼酸能够积累并且在脱氢奎尼酸脱水酶的催化下经过3-脱氢莽草酸合成莽草酸。Chen等在ydiB基因完全缺失的大肠杆菌SA4中过量表达莽草酸脱氢酶基因,以减少代谢流中3-脱氢奎尼酸向奎尼酸的转化,促进3-脱氢莽草酸的积累,进而促进莽草酸的生物合成[20]。有研究者将 1 mmol/L 甲基-α-D-葡萄糖吡喃糖苷加到葡萄糖限制性培养基中作为碳源,经过48 h的发酵培养,副产物奎尼酸的产量由 19 g/L 减少到2.8 g/L,目的产物莽草酸的产量从原来的 28 g/L 升高到35 g/L,莽草酸的产率由15%上升至19%。该研究表明,以1 mmol/L甲基-α-D-葡萄糖吡喃糖苷作为碳源可以明显提高莽草酸的合成量,同时减少副产物奎尼酸的产生[6]。在烟草中,存在同时具有3-脱氢奎尼酸脱水酶(aroD)和莽草酸脱氢酶(aroE)催化活性的双功能酶,由aroD*E基因编码。将烟草来源的aroD*E基因导入aroD、aroE基因同时被敲除的重组菌中,经过66 h的发酵培养,莽草酸的产量达到34 g/L,产率达到15%。发酵液中莽草酸、奎尼酸、3-脱氢莽草酸的物质的量的比为 29 ∶ 1.0 ∶ 5.7,可见副产物奎尼酸的产量明显减少。
6 辅因子烟酰胺腺嘌呤二核苷酸磷酸(NADPH)的表达
辅因子是一类能够使底物与酶活性部位结合或者使酶更具有活性的金属离子或辅酶。有些氧化还原酶催化的反应是需要以还原态烟酰胺腺嘌呤二核苷酸(NADH)、NADPH为辅酶的,氧化还原力的不平衡,不仅会影响细胞的连续生长、目的产物的合成,还会导致代谢碳流从需要生成的目的产物的代谢途径流转向其他途径而产生副产物。因此,控制辅因子的生物合成供应对于进一步提高目的产物产量非常重要[28-29]。研究表明,维持NADPH的可用浓度可以提高代谢产物的产量。Kabus等在谷氨酸棒状杆菌体内过表达来自大肠杆菌的转氢酶基因pntAB发现,在以葡萄糖为碳源的基本培养基中经过72 h的发酵培养,赖氨酸的浓度从474 mmol/L升高到了522 mmol/L[30]。在莽草酸代谢途径中,莽草酸脱氢酶需要NADPH辅因子的作用才能催化 3- 脱氢莽草酸转化为莽草酸。目前,多数莽草酸工程菌的构建都会过表达关键酶基因aroE,这样就会出现NADPH的还原力供应不足。在大肠杆菌体内,转氢酶(membrane-bound transhydrogenase,简称PntAB)和烟酰胺腺嘌呤二核苷酸(NAD)激酶(NAD kinase,简称NADK)可以催化NADH与NADPH之间相互转化,因此可以强化基因pntAB、nadK的表达,缓解还原力不平衡的问题。Cui等对NADPH对莽草酸途径中莽草酸产量的影响进行研究,他们将pntAB、nadK基因分别整合到大肠杆菌染色体中,以增加细胞内NADPH的供应量,摇瓶培养结果显示,整合nadK基因后的重组菌SA116产率达到33%,产量为3.12 g/L[31]。对于戊糖磷酸途径,可以过表达戊糖磷酸途径中的NADPH代谢关键酶,例如葡萄糖-6-磷酸脱氢酶(glucose-6-phosphate-1-dehydrogenase,由zwf基因编码)和6-磷酸葡萄糖酸脱氢酶(6-phosphogluconate dehydrogenase,由gnd基因编码)。Rodriguez等敲除大肠杆菌的PTS系统和pykF基因,同时过表达aroGfrb、aroE、aroB、aroD、tktA和zwf基因,分批发酵结果显示,重组工程菌AR36产生莽草酸的终浓度为43.4 g/L,产率为42%[23],这是目前研究莽草酸工程菌株中获得的最高产率。
7 引导代谢平衡的模块优化策略
莽草酸途径是一个较为复杂的代谢途径,有研究发现,通过过量表达上游代谢途径关键酶基因而使上游代谢碳流急剧增加时,由于下游代谢途径无法正常消耗强大的上游碳代谢流,会造成中间产物的过量积累,过量存在的中间产物会对细胞内的代谢途径产生反馈抑制,或者产生细胞生长不需要的副产物,从而影响菌体的正常生长。因此,Wu等引入了模块化代谢工程改造理论,系统改造各個模块的碳代谢流量,以期找到一条最优的途径平衡方式,使代谢流更多地从可再生的廉价底物流向高附加值的目的化合物;Wu等引入模块化代谢工程改造理论优化黄酮骨架物质的合成途径,首先将总途径分为4个模块,通过系统改变各个模块的代谢通量来寻找途径中的瓶颈步骤,通过4个不同拷贝数的质粒和2个不同强度的启动子来改变模块的代谢通量以平衡合成途径,经过多轮模块优化后,生松素、柚皮素的产量分别提高至84.2、105.1 mg/L[32]。Zhang等在C. glutamicum中引入来源于古细菌Mcc. maripaludis和Msc. mazei的DKFP(6-deoxy-5-ketofructose 1-phosphate,即6-脱氧-5-酮果糖1-磷酸)途径基因,以1个丙酮酸合酶基因、2个DKFP合酶基因、2个ADTH合酶基因、1个DHQ合酶基因为元件,通过不同组合方式组装了3组模块,结果表明,同时表达异源莽草酸代谢途径与自身莽草酸代谢途径的重组谷氨酸棒状杆菌RES167ΔaroK/pDKFP-3比没有导入异源代谢途径的原始菌株RES167ΔaroK的莽草酸产量提高了57%,莽草酸产量从0.77 g/L提高到了1.20 g/L[11]。研究者从合成生物学的角度出发,通过一系列代谢途径优化、基因元件筛选与模块组合,完成了对莽草酸代谢途径的异源装配,进一步对莽草酸代谢途径模块优化策略实施了验证。
8 总结与展望
通过对莽草酸代谢途径的改造,包括解除莽草酸代谢途径中的反馈抑制、过表达莽草酸代谢途径中的关键酶基因、代谢途径中NADPH辅因子再生等,结果发现,经过遗传改造的基因工程菌株合成莽草酸的产量有明显提升。莽草酸是合成芳香族类氨基酸(L-苯丙氨酸、L-酪氨酸、L-色氨酸)的重要前体化合物,同时因其自身的药用价值,具有广阔的市场前景与经济价值。以葡萄糖为起始合成原料,通过基因改造工程菌生产莽草酸的方法,具有发酵周期短、可控性强、可持续性发展且对环境友好等特点,非常具有发展潜力与应用价值。尽管基因改造的工程菌较野生型的莽草酸发酵产量有所提高,但是仍然存在进一步提高的空间。今后的研究可以从以下几个方面考虑:
(1)通过定点突变解除DAHP合酶的反馈抑制后,可以通过多级质谱或者核磁共振技术分析细胞全局的代谢流,从而找出细胞内可能存在的其他限制性步骤。
(2)通过基因过表达的方式提高了辅因子NADPH的供应量,进一步可以研究如何提高莽草酸代谢途径中所需要的其他辅因子,例如ATP和丙二酰辅酶A等。
(3)研究表明,以葡萄糖为碳源进行发酵培养,菌株生长较慢,但是莽草酸生产持续时间较长;以甘油为碳源时,菌株生长较快,但是莽草酸合成过早停止,因此可以尝试葡萄糖结构类似物或者甘油结构类似物,观察发酵结果是否能达到生产速率与持续生产时间的正相关。
(4)可以考虑构建多种功能的菌株进行联合培养发酵,例如构建特异性消耗副产物的菌株,与莽草酸菌株一起培养,以改善莽草酸生产菌株的生长环境,提高莽草酸的产量。
参考文献:
[1]Ma Y,Sun J N,Xu Q P,et al. Inhibitory effects of shikimic acid on platelet aggragation and blood coagulation[J]. Acta Pharmaceutica Sinica,2000,5(5):600-612.
[2]Abbott A. Avian flu special:whats in the medicine cabinet?[J]. Nature,2005,435(7041):407-409.
[3]Herrmann K M. The shikimate pathway:early steps in the biosynthesis of aromatic compounds[J]. Plant Cell,1995,7(7):907-919.
[4]Richards T A,Dacks J B,Campbell S A,et al. Evolutionary origins of the eukaryotic shikimate pathway:gene fusions,horizontal gene transfer,and endosymbiotic replacements[J]. Eukaryotic Cell,2006,5(9):1517.
[5]Johansson L,Lindskog A,Silfversparre G,et al. Shikimic acid production by a modified strain of E. coli (W3110.shik1) under phosphate-limited and carbon-limited conditions[J]. Biotechnology & Bioengineering,2010,92(5):541-552.
[6]Knop D R,Draths K M,Chandran S S,et al. Hydroaromatic equilibration during biosynthesis of shikimic acid[J]. Journal of the American Chemical Society,2001,123(42):10173-10182.
[7]汪 華,崔志峰. 莽草酸生物合成途径的调控[J]. 生物技术通报,2009(3):50-53.
[8]Cortés-Tolalpa L, Gutiérrez-Ríos R M, Martínez L M,et al. Global transcriptomic analysis of an engineered Escherichia coli strain lacking the phosphoenolpyruvate:carbohydrate phosphotransferase system during shikimic acid production in rich culture medium[J]. Microbial Cell Factories,2014,13(1):28.
[9]Liconacassani C,Lara A R,Cabreravalladares N,et al. Inactivation of pyruvate kinase or the phosphoenolpyruvate:sugar phosphotransferase system increases shikimic and dehydroshikimic acid yields from glucose in Bacillus subtilis[J]. Journal of Molecular Microbiology & Biotechnology,2014,24(1):37.
[10]Zhang B,Zhou N,Liu Y M,et al. Ribosome binding site libraries and pathway modules for shikimic acid synthesis with Corynebacterium glutamicum[J]. Microbial Cell Factories,2015,14(1):1-14.
[11]Zhang B,Jiang C Y,Liu Y M,et al. Engineering of a hybrid route to enhance shikimic acid production in Corynebacterium glutamicum[J]. Biotechnology Letters,2015,37(9):1861-1868.
[12]Rawat G,Tripathi P,Yadav S,et al. A natural isolate producing shikimic acid:isolation,identification,and culture condition optimization[J]. Applied Biochemistry & Biotechnology,2013,169(8):2290.
[13]Rawat G,Tripathi P,Yadav S,et al. An interactive study of influential parameters for shikimic acid production using statistical approach,scale up and its inhibitory action on different lipases[J]. Bioresource Technology,2013,144(6):675-679.
[14]Tripathi P,Rawat G,Yadav S,et al. Fermentative production of shikimic acid:a paradigm shift of production concept from plant route to microbial route[J]. Bioprocess & Biosystems Engineering,2013,36(11):1665-1673.
[15]Tripathi P,Rawat G,Yadav S,et al. Shikimic acid,a base compound for the formulation of swine/avian flu drug:statistical optimization,fed-batch and scale up studies along with its application as an antibacterial agent[J]. Antonie van Leeuwenhoek,2015,107(2):419-431.
[16]Ghosh S,Banerjee U C. Generation of aroE overexpression mutant of Bacillus megaterium for the production of shikimic acid[J]. Microbial Cell Factories,2015,14(1):69.
[17]Liu D F,Ai G M,Zheng Q X,et al. metabolic flux responses to genetic modification for shikimic acid production by Bacillus subtilis strains[J]. Microbial Cell Factories,2014,13(1):1-11.
[18]刘东风. 产莽草酸枯草芽孢胞杆菌代谢工程改造及代谢流分析[D]. 合肥:中国科学技术大学,2014.
[19]Chen K,Dou J,Tang S H,et al. Deletion of the aroK gene is essential for high shikimic acid accumulation through the shikimate pathway in E. coli[J]. Bioresource Technology,2012,119(9):141-147.
[20]Chen X Z,Li M M,Zhou L,et al. metabolic engineering of Escherichia coli for improving shikimate synthesis from glucose[J]. Bioresource Technology,2014,166(8):64-71.
[21]Chandran S S,Yi J,Draths K M,et al. Phosphoenolpyruvate availability and the biosynthesis of shikimic acid[J]. Biotechnology Progress,2003,19(3):808.
[22]肖夢榕. 改进大肠杆菌PTS系统构建莽草酸生产菌[D]. 无锡:江南大学,2014.
[23]Rodriguez A,Martinez J A,Baez-Viveros J L,et al. Constitutive expression of selected genes from the pentose phosphate and aromatic pathways increases the shikimic acid yield in high-glucose batch cultures of an Escherichia coli strain lacking PTS and pykF[J]. Microbial Cell Factories,2013,12(1):86.
[24]Bongaerts J,Kramer M,Mulle U,et al. metabolic engineering for microbial production of aromatic amino acids and derived compounds[J]. metabolic Engineering,2001,3(4):289-300.
[25]仲 楠. 产莽草酸基因工程菌的构建及发酵研究[D]. 北京:北京化工大学,2012.
[26]Draths K M,Knop D R,Frost J W. Shikimic acid and quinic acid:replacing isolation from plant sources with recombinant microbial biocatalysis[J]. Journal of the American Chemical Society,1999,121(7):1603-1604.
[27]Yi J,Draths K M,Li K,et al. Altered glucose transport and shikimate pathway product yields in E. coli[J]. Biotechnology Progress,2010,19(5):1450-1459.
[28]Fumio M,Jun I,Takashi K,et al. Increased isobutanol production in Saccharomyces cerevisiae by eliminating competing pathways and resolving cofactor imbalance[J]. Microbial cell factories,2013,12(1):119.
[29]Shi A,Zhu X,Lu J,et al. Activating transhydrogenase and NAD kinase in combination for improving isobutanol production[J]. metabolic Engineering,2013,16(16):1-10.
[30]Kabus A,Georgi T,Wendisch V F,et al. expression of the Escherichia coli pntAB genes encoding a membrane-bound transhydrogenase in Corynebacterium glutamicum improves l-lysine formation[J]. Applied Microbiology & Biotechnology,2007,75(1):47-53.
[31]Cui Y Y,Chen L,Zhang Y Y,et al. Production of shikimic acid from Escherichia coli through chemically inducible chromosomal evolution and cofactor metabolic engineering[J]. Microbial Cell Factories,2014,13(1):21.
[32]Wu J J,Du G C,Zhou J W,et al. metabolic engineering of Escherichia coli for (2S)-pinocembrin production from glucose by a modular metabolic strategy[J]. metabolic Engineering,2013,16(1):48-55. 王 森,唐 蛟,王晓森,等. 加气灌溉对土壤、作物生长和水肥利用的影响综述[J]. 江苏农业科学,2019,47(7):24-27.




 加载中...
加载中...