


 13720223167
13720223167



(华中农业大学 食品科学技术学院,湖北 武汉,430070)
摘 要 S-腺苷甲硫氨酸(S-adenosylmethionine,SAM),可作为营养补充剂用于改善人体健康,对人体新陈代谢至关重要。在解淀粉芽胞杆菌从头合成SAM的过程中,脂肽类表面活性剂常导致能量损失、发酵起泡等问题,不利于SAM的积累。通过研究敲除解淀粉芽孢杆菌脂肽类表面活性剂合成基因ituD和srfAC,探究该类基因对SAM从头合成的影响。实验结果显示:单敲除ituD基因并未对菌体生长和SAM合成产生显著的影响,单敲除srfAC基因促进SAM产量提高了23%,对菌体生长影响不显著;叠加敲除ituD和srfAC后,SAM产量提高32%,且对菌体生长没有显著性影响。研究首次证实敲除脂肽类表面活性剂基因可显著提高SAM产量,研究结果为SAM的代谢工程育种提供了新策略。
关键词 S-腺苷甲硫氨酸;解淀粉芽胞杆菌;脂肽类表面活性剂;基因敲除;代谢工程
S-腺苷甲硫氨酸(S-adenosylmethionine,SAM),是生物体内含硫氨基酸代谢的一个关键组成部分,由甲硫氨酸和三磷酸腺苷通过蛋氨酸腺苷转移酶合成[1]。SAM在细胞中主要参与转甲基化[2]、转硫化[3]和脂肪多胺合成[4],被视为生物体中的关键代谢物,常用于预防和改善肝病、关节炎、抑郁症、阿尔茨海默氏症等多种疾病[5-7]。欧美国家已将其批准用作膳食补充剂[8]。因此,开发SAM相关产品具有重要的意义。
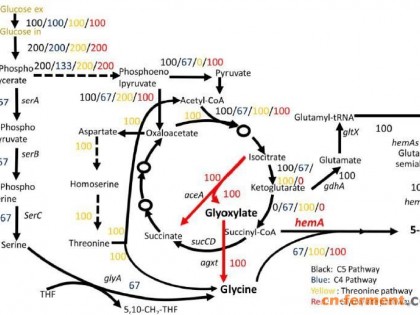
微生物发酵法是生产SAM的主要方法。早期的研究普遍通过添加大量甲硫氨酸前体物来实现SAM的高效合成,产量可达10 g/L以上[9],但甲硫氨酸添加量大、成本高,且转化率低,导致SAM价格昂贵[10]。进而有研究者以廉价糖类碳源从头合成SAM,以期降低发酵成本[11]。然而,目前从头合成SAM的产量仍较低,尚需开展深入研究。课题组前期通过解淀粉芽胞杆菌从头合成SAM,通过途径耦合代谢工程策略强化了SAM的合成[12],该法在SAM的从头合成中具有重要的应用潜力,但仍需开展大量研究工作。
解淀粉芽胞杆菌可大量合成两亲性脂肽类表面活性剂副产物[13],不仅消耗大量的能量,而且这些表面活性剂可导致发酵过程起泡严重,影响发酵过程[14]。芽胞杆菌产生的脂肽类表面活性剂主要包括iturin和surfactin,其中iturin主要由ituDABC操纵子基因编码[15-16],surfactin主要由srfA-ABCD操纵子基因编码[17-18]。通过基因敲除技术阻断iturin和surfactin的合成有望促进SAM的合成,然而目前还未见相关报道。因此,本研究通过基因敲除技术敲除解淀粉芽孢杆菌中的ituD和SrfAC基因,阻断iturin和surfactin的合成,以期降低发酵过程脂肽类副产物和发酵泡沫,以期提高SAM产量。
1 材料与方法
1.1 材料与试剂
1.1.1 菌株和质粒
本研究所用到的菌株和质粒如表1和表2所示。其中大肠杆菌DH5α用于构建载体。
表1 实验用菌株
Table 1 Strains used in this study

表2 实验用质粒
Table 2 Plasmids used in this study
注:Tetr:四环素抗性;Kanr:卡那霉素抗性;orits:温敏型复制子
1.1.2 PCR引物
本研究所用的主要引物见表3。
表3 本研究所使用的PCR引物
Table 3 Primers used for PCR in this study
注:粗体为限制性内切酶酶切位点,划线为重叠延伸PCR (SOE-PCR)的重叠区域
1.1.3 主要培养基
SAM发酵培养基(g/L):蔗糖80,蛋白胨10,玉米浆5,天冬氨酸3,尿素2,NaCl 2.5,(NH4)2SO4 6.3,KH2PO4 3,MgSO4·7H2O 4.2,pH 6.5。
LB培养基(g/L):蛋白胨10.0,酵母浸出粉5.0,NaCl 10.0,pH 7.2,固体培养基加琼脂1.5%。
1.1.4 工具酶和试剂
DNA抽提试剂盒,武汉楚诚正茂科技工程有限公司;TransStartR FastPfu DNA聚合酶和TransStartR easyTaq DNA聚合酶,北京全式金公司;DNA限制性内切酶、T4 DNA连接酶,DNA回收试剂盒和质粒抽提试剂盒,美国OmegaBio-Tek公司;DL5000 Marker,TaKaRa公司;琼脂糖,Spanish公司;Kan、Tet、溶菌酶、山梨醇、甘露醇等试剂等均为进口分装,武汉楚诚正茂科技工程有限公司,biosharp;甲醇为色谱级,其他主要生化试剂均为国产分析级纯或进口分装。
1.1.5 仪器与设备
Agilent Technologies 1260高效液相色谱,美国Agilent公司;Thermal Cycler (My Cycler) PCR仪,美国 Bio-Rad 公司;DYY-8C型电泳仪,北京六一仪器厂;CR21G高速冷冻离心机,日本Hitachi公司;HQL300B恒温大幅振荡摇床,武汉中科科仪技术发展有限责任公司;SKP-02.420电热恒温培养箱,黄石市恒丰医疗器械有限公司;Legend Micro17R高速冷冻离心机,美国Thermo Fisher Scientific 公司。
1.2 实验方法
1.2.1 敲除载体的构建
以ituD基因为例,选取B. amyloliquefaciens HZ-12的基因组DNA作为模板,利用上游同源臂引物(ΔituD-AF, ΔituD-AR)和下游同源臂引物(ΔituD-BF,ΔituD-BR)分别扩增出同源臂序列A和B,产物纯化回收后通过SOE-PCR用引物ΔituD-AF、ΔituD-BR将A和B连接起来,并与T2(2)-ori载体同时用限制性内切酶SpeI和XbaI进行双酶切,回收后用T4 DNA连接酶连接,采用CaCl2转化法转化到E.coli DH5α感受态细胞。钙转化成功后提取质粒进行双酶切验证并送往测序公司测序,构建成功的敲除质粒命名为T2(2)-oriΔituD。
1.2.2 基因敲除菌株的构建[12]
以ituD基因为例,将构建成功的质粒电转化入HSAM4感受态细胞中,涂布至含Kan的抗性平板,37 ℃静置培养16~18 h。挑选转化子单菌落于Kan平板上划线,培养8~12 h后进行PCR菌落验证(引物T2-F,T2-R)。将电转正确的菌株在45 ℃下进行单交换,ΔituD-YF/YR引物和T2引物验证;获得的单交换菌株在37 ℃下进行双交换,挑选在LB板上长而在Kan抗性板上不长的单菌落,用ΔituD-YF和ΔituD-YR验证,筛选到敲除菌株,命名为HSAM4ΔituD。
1.2.3 发酵培养条件
从超低温冰箱中取出保藏的菌种划线LB平板培养基中,37 ℃培养基12 h。挑取适量菌体于装有50 mL LB液体培养基的250 mL三角瓶中,37 ℃,180 r/min培养9 h。吸取体积分数1.5%的菌液接种于装有25 mL发酵培养基的250 mL三角瓶中,37 ℃、180 r/min发酵培养,每个菌设置3个摇瓶重复。
1.2.4 SAM液相检测[12]
取发酵液0.5 mL,加1.5 mL 0.4 mol/L高氯酸,提取1 h(每15 min振荡1次),10 000 r/min离心5 min。取800 μL上清液,加100 μL,2 mol/L NaOH调pH。利用HPLC检测,色谱柱为XDB-C18(规格:5 μm,4.6 mm×150 mm),检测器为紫外检测器,流动相为V(甲醇)∶V(40 mmol/L NH4H2PO4 2 mmol/L庚烷磺酸钠)=18∶82,流速为0.8 mL/min,进样量为20 μL,柱温30 ℃,检测波长为254 nm。
1.2.5 数据统计分析
每组实验设计3个重复,采用SPSS 20.0进行数据统计分析,评估数据差异的显著性,计算出平均值和标准偏差,采用Origin 8.5进行图表绘制。
2 结果与分析
2.1 ituD基因缺失对解淀粉芽孢杆菌合成SAM的影响
2.1.1 ituD基因缺失菌株的构建
利用1.2.1小节中所述的方法构建重组载体T2(2)-oriΔituD,利用表3中相应的引物ΔituD-AF/AR,ΔituD-BF/BR扩增用于构建T2(2)-oriΔituD的上下游同源臂。敲除ituD基因的上下游同源臂大小分别为513和504 bp,上下游同源臂经SOE-PCR融合后的产物大小为1 017 bp,上下游同源臂SOE-PCR融合片段与T2(2)-ori质粒经酶切和酶连后转化至E.coli DH5α,选取验证正确的阳性转化子,提取质粒并且测序结果正确无误,表明敲除质粒T2(2)-oriΔituD构建成功。
将已构建完成的重组质粒T2(2)-oriΔituD电转化至HASM4中,筛选出阳性转化子,并按照1.2.2的方法进行单交换和双交换筛选突变株。经菌落PCR验证正确的双交换菌株,提取基因组DNA,并利用表3中相应的引物ΔituD-YF/YR验证原始菌株HSAM4和阳性交换突变株,片段大小分别为2 508、1 290 bp,两片段相差的长度正好是ituD基因的长度,PCR验证电泳图如图1所示。且纯化回收阳性交换突变株PCR产物,经测序确认碱基正确无误,说明缺失ituD基因的菌株HSAM4ΔituD构建成功。
1-ΔituD-YF 和ΔituD-YR PCR验证HSAM4(2 508 bp);
2-ΔituD-YF 和ΔituD-YR PCR验证HSAM4ΔituD(1 290 bp)
图1 HSAM4ΔituD菌株的PCR验证电泳图
Fig.1 Electrophoretogram of PCR validation for
HSAM4ΔituD
2.1.2 ituD基因缺失对SAM合成的影响
为了考察ituD基因的缺失对B. amyloliquefaciens HSAM4合成SAM的影响,按照1.1.3和1.2.3小节的发酵培养基和培养条件对工程菌株HSAM4ΔituD进行发酵验证,发酵周期为60 h。发酵结束后取样检测SAM产量和菌体生物量(OD600),结果如图2所示。
图2 缺失ituD基因对SAM合成的影响
Fig.2 Effect of ituD knockout on SAM fermentation
由图2可以看出,原始菌 HSAM4与HSAM4ΔituD的生物量无显著差异,说明ituD基因缺失对菌体生长无影响。此外,缺失菌株HSAM4ΔituD的SAM产量与原始菌 HSAM4相比,没有显著性提高。上述结果说明敲除ituD基因对菌体生长和产物的合成未产生显著性影响。
2.2 srfAC基因缺失对解淀粉芽孢杆菌合成SAM的影响
2.2.1 srfAC基因缺失菌株的构建
敲除质粒T2(2)-oriΔsrfAC来自本实验室保存。将T2(2)-oriΔsrfAC电转化至HASM4中,筛选出阳性转化子,并按照1.2.2的方法进行单交换和双交换筛选突变株。经菌落PCR验证正确的双交换菌株,提取基因组DNA,并利用表3中相应的引物ΔsrfAC-YF/YR验证原始菌株HSAM4和阳性交换突变株,片段大小分别为 2 704和1 657 bp,两片段相差的长度正好是srfAC基因的长度,PCR验证电泳图如图3所示。纯化回收阳性交换突变株PCR产物,经测序确认碱基正确无误,说明缺失srfAC基因的菌株HSAM4ΔsrfAC构建成功。
1-ΔsrfAC-YF 和ΔsrfAC-YR PCR验证HSAM4(2 704 bp);
2-ΔsrfAC-YF 和ΔsrfAC-YR PCR验证HSAM4ΔsrfAC(1 657 bp)
图3 HSAM4ΔsrfAC菌株的PCR验证电泳图
Fig.3 Electrophoretogram of PCR validation for
HSAM4ΔsrfAC
2.2.2 srfAC 基因缺失对SAM合成的影响
为了考察 srfAC 基因的缺失对B. amyloliquefaciens HSAM4合成SAM的影响,按照1.1.3和1.2.3小节的发酵培养基和培养条件对工程菌株HSAM4ΔsrfAC进行发酵验证,发酵周期为60 h。发酵结束后取样检测SAM产量和菌体生物量(OD600),结果如图4所示。
图4 缺失srfAC基因对SAM合成的影响
Fig.4 Effect of srfAC knockout on SAM fermentation
由图4可以看出,工程菌 HSAM4ΔsrfAC的生物量相比于原始菌没有发生显著性变化。缺失菌株 HSAM4ΔsrfAC的SAM 产量为 208.24 mg/L,比原始菌HSAM4提高了23%,差异显著。推测其原因,srfAC基因编码surfactin合成酶[17],影响脂肽的环化与释放,减少了表面活性剂surfactin的合成,可降低副产物和发酵产泡现象,从而促进SAM的合成。
2.3 ituD基因和srfAC基因双缺失对解淀粉芽孢杆菌合成SAM的影响
2.3.1 ituD基因和srfAC基因双缺失菌株的构建
将T2(2)-oriΔsrfAC电转化至HASM4ΔituD中,筛选出阳性转化子,并按照1.2.2的方法进行单交换和双交换筛选突变株。经菌落PCR验证正确的双交换菌株,提取基因组DNA,并利用表3中相应的引物ΔituD-YF/YR和ΔsrfAC-YF/YR验证原始菌株HSAM4和阳性交换突变株,与原始菌相比分别相差ituD基因和srfAC基因的长度,PCR验证电泳图如图5所示。纯化回收阳性交换突变株PCR产物,经测序确认碱基正确无误,说明缺失ituD和srfAC基因的菌株HSAM4ΔituDΔsrfAC构建成功。
1-ΔituD-YF 和ΔituD-YR PCR验证HSAM4(2 508 bp);
2-ΔituD-YF 和ΔituD-YR PCR验证HSAM4ΔituDΔsrfAC(1 290 bp);
3-ΔsrfAC-YF 和ΔsrfAC-YR PCR验证HSAM4(2 704 bp);
4-ΔsrfAC-YF 和ΔsrfAC-YR PCR
验证HSAM4ΔituDΔsrfAC(1 657 bp)
图5 HSAM4ΔituDΔsrfAC菌株的PCR验证电泳图
Fig.5 Electrophoretogram of PCR validation for
HSAM4ΔituDΔsrfAC
2.3.2 ituD基因和srfAC基因双缺失对SAM合成的影响
按照1.1.3和1.2.3小节的发酵培养基和培养条件对工程菌株HSAM4ΔituDΔsrfAC进行发酵验证,发酵周期为60 h。发酵结束后取样检测 SAM 产量和菌体生物量(OD600),结果如图6所示。
图6 缺失ituD,srfAC基因对SAM合成的影响
Fig.6 Effect of ituD,srfAC knockout on SAM
fermentation
由图6可以看出,工程菌HSAM4ΔituDΔsrfAC的生物量相比于原始菌 HSAM4无明显变化,说明 ituD和srfAC基因双缺失对菌体的生长无影响。缺失菌株HSAM4ΔituDΔsrfAC的SAM的产量达222.95 mg/L,相比于原始菌 HSAM4提高了32%。这可能是敲除ituD基因和srfAC基因,同时阻断了iturin和surfactin的合成[15,17],降低发酵副产物和发酵产泡现象,进一步提高了SAM产量。
3 结论
S-腺苷甲硫氨酸是存在于生物体中的必需功能因子[19]。在不添加甲硫氨酸的前提下,通过廉价碳源实现SAM的高效从头合成具有重要的意义。解淀粉芽胞杆菌在从头合成SAM的过程中显示了初步的效果[12],为有效提高SAM的从头合成产量,本文基于基因敲除技术,成功构建了ituD 基因缺失菌株HSAM4ΔituD,srfAC 基因缺失菌株HSAM4ΔsrfAC,ituD、srfAC基因双缺失菌株 HSAM4ΔituDΔsrfAC。SAM发酵结果显示,缺失ituD、srfAC基因对菌株生长几乎无影响,双缺失菌株HSAM4ΔituDΔsrfAC的SAM产量可提高至原始菌株HSAM4的1.32倍。
ZHANG等[20]研究发现,利用Bacillus subtilis ATCC 6051a发酵生产环糊精糖基转移酶过程中,敲除该菌株的srfAC基因可有效控制发酵过程中的泡沫。Iturin和surfactin作为两亲性脂肽类表面活性物质,是液体深层发酵产泡的重要原因。由此推测,本研究ituD和srfAC基因的缺失,阻断了iturin和surfactin的产生,不仅可减少发酵过程中副产物的生成及不必要的能量消耗,而且可通过降低表面活性剂改善发酵产泡现象,这可能是SAM产量提高的主要原因。本研究为SAM从头合成的代谢工程育种提供了新的思路和方向。
参考文献
[1] PANZA F, FRISARDI V, CAPURSO C, et al. Polyunsaturated fatty acid and S-adenosylmethionine supplementation in predementia syndromes and Alzheimer's disease: a review[J]. The Scientific World Journal, 2009, 9: 373-389.
[2] KRUGLOVA M P, GRACHEV S V, BULGAKOVA P O, et al. Low S-adenosylmethionine/S-adenosylhomocysteine ratio in urine is associated with chronic kidney disease[J]. Laboratory Medicine, 2020, 51(1): 80-85.
[3] SINNER E K, LICHSTRAHL M S, LI R, et al. Methylations in complex carbapenem biosynthesis are catalyzed by a single cobalamin-dependent radical S-adenosylmethionine enzyme[J]. Chemical Communications, 2019, 55(99): 14 934-14 937.
[4] CAMPonESCHI F, MUZZIOLI R, CIOFI-BAFFONI S, et al. Paramagnetic 1H NMR spectroscopy to investigate the catalytic mechanism of radical S-adenosylmethionine enzymes[J]. Journal of Molecular Biology, 2019, 431(22): 4 514-4 522.
[5] RAIKHELSON K L, KonDRASHINA E A. Аdеmethionine in the treatment of fatigue in liver diseases: a systematic review[J]. Terapevticheskii Arkhiv, 2019, 91(2): 134-142.
[6] LANDGRAF B J, MCCARTHY E L, BOOKER S J. Radical S-adenosylmethionine enzymes in human health and disease[J]. Annual Review of Biochemistry, 2016, 85: 485-514.
[7] MARTNEZ-UA M, VARELA-REY M, MESTRE D, et al. S-Adenosylmethionine increases circulating very-low density lipoprotein clearance in non-alcoholic fatty liver disease[J]. Journal of Hepatology, 2015, 62(3): 673-681.
[8] MATO J M, MARTNEZ-CHANTAR M L, LU S C. S-adenosylmethionine metabolism and liver disease[J]. Annals of Hepatology, 2015, 12(2): 183-189.
[9] KANAI M, MIZUNUMA M, FUJII T, et al. A genetic method to enhance the accumulation of S-adenosylmethionine in yeast[J]. Applied Microbiology and Biotechnology, 2017, 101(4): 1 351-1 357.
[10] CHU J, QIAN J, ZHUANG Y, et al. Progress in the research of S-adenosyl-L-methionine production[J]. Applied Microbiology and Biotechnology, 2013, 97(1): 41-49.
[11] CHEN Y, XU D, FAN L, et al. Manipulating multi-system of NADPH regulation in Escherichia coli for enhanced S-adenosylmethionine production[J]. RSC Advances, 2015, 5(51): 41 103-41 111.
[12] RUAN L, LI L, ZOU D, et al. metabolic engineering of Bacillus amyloliquefaciens for enhanced production of S-adenosylmethionine by coupling of an engineered S-adenosylmethionine pathway and the tricarboxylic acid cycle[J]. Biotechnology for Biofuels, 2019, 12(1): 211.
[13] 卢彩鸽, 张殿朋, 刘霆, 等. 解淀粉芽胞杆菌 MH71 的生防活性及脂肽类抗生素基因检测[J].植物保护, 2015, 41(3): 12-18.
[14] HOEKS F W, VAN WEES-TANGERMAN C, LUYBEN K C A M, et al. Stirring as foam disruption (SAFD) technique in fermentation processes[J]. The Canadian Journal of Chemical Engineering, 1997, 75(6): 1 018-1 029.
[15] 郭丽清. 苏云金芽孢杆菌中脂肽类抗生素 Iturins 的研究[D].福州:福建农林大学, 2010.
[16] LEES F L, VELHO R V, CALDAS D G G, et al. Influence of pH and temperature on the expression of sboA and ituD genes in Bacillus sp. P11[J]. Antonie Van Leeuwenhoek, 2013, 104(1): 149-154.
[17] 刘丽霞. Surfactin合成酶相关基因研究[D].南京:南京农业大学, 2012.
[18] WANG X, CHEN Z, FENG H, et al. Genetic variants of the oppA gene are involved in metabolic regulation of surfactin in Bacillus subtilis[J]. Microbial Cell Factories, 2019, 18(1): 141.
[19] YAN L, LIANG X, HUANG H, et al. S-adenosylmethionine affects cell cycle pathways and suppresses proliferation in liver cells[J]. Journal of Cancer, 2019, 10(18): 4 368-4 379.
[20] ZHANG K, DUAN X, WU J. Multigene disruption in undomesticated Bacillus subtilis ATCC 6051a using the CRISPR/Cas9 system[J]. Scientific Reports, 2016, 6(1): 1-11.
Enhancement of S-adenosylmethionine synthesis in Bacillus amyloliquefaciensdeleted surfactant synthesis
JIANG Cong, GUO Ailing, WEI Xuetuan*
(College of Food Science and Technology, Huazhong Agricultural University, Wuhan 430070, China)
ABSTRACT S-Adenosylmethionine (SAM) is highly important for human metabolism, and it can be used as a nutritional supplement to improve human health. During the de novo synthesis of SAM by Bacillus amyloliquefaciens, lipopeptide surfactants usually lead to energy loss and fermentation foaming, which may hinder the accumulation of SAM. In this study, the lipopeptide surfactants synthesis genes of ituD and srfAC were deleted and effects of these genes on SAM synthesis were investigated. The fermentation results showed that deletion of ituD had no significant effect on cell growth and SAM synthesis. On the other hand, deletion of srfAC promoted the SAM titer by 23%, and no significant effect was found on the cell growth. After double mutation of ituD and srfAC, the SAM titer was increased by 32%, and the cell growth was not affected significantly. This study confirms that blocked lipopeptide surfactants synthesis can significantly enhance the SAM production, which provides a new strategy for metabolic engineering breeding of SAM.
Key words S-adenosylmethionine; Bacillus amyloliquefaciens; lipopeptide surfactant; gene knockout; metabolic engineering
DOI:10.13995/j.cnki.11-1802/ts.023690
第一作者:硕士研究生(魏雪团副教授为通讯作者,E-mail:weixuetuan@mail.hzau.edu.cn)
基金项目:国家重点研发计划(2019YFA0906400)
收稿日期:2020-02-19,改回日期:2020-02-27
引用格式:姜聪,郭爱玲,魏雪团.通过敲除解淀粉芽胞杆菌表面活性剂基因促进S-腺苷甲硫氨酸合成[J].食品与发酵工业,2020,46(11):17-22.JIANG Cong, GUO Ailing, WEI Xuetuan. Enhancement of S-adenosylmethionine synthesis in Bacillus amyloliquefaciens deleted surfactant synthesis[J].Food and Fermentation Industries,2020,46(11):17-22.




 加载中...
加载中...