


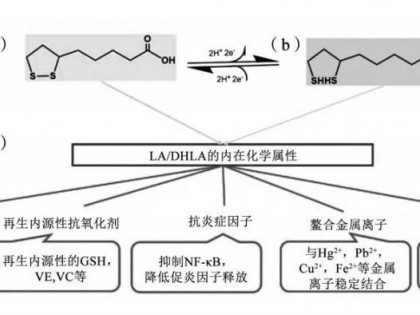
 13720223167
13720223167



酿造酵母生长繁殖需要非常复杂的麦汁环境,这其中包括不同含量的多种营养物质。在麦汁中,酵母代谢所需的主要氮源是由麦芽蛋白分解形成的单体氨基酸,小分子肤和钱离子。酿造麦汁中包含20种基本氨基酸中的19种,与麦芽糖一样,氨基酸的降解是有序的。氨基酸按照酵母同化的方式可分为四组(见表1)。这种分类方式也可能因酿造菌株或贮藏酵母的种类而略有差异。麦汁中的单体氨基酸和小分子肤(二肤和三肤)可以通过游离氨基酸(FAN)定量。FAN是一项表明潜在的酵母生长和发酵力的重要指标。

材料和方法
.发酵
在已发酵的2L圆筒中进行静态发酵,用橡皮塞和发酵环密封,其中含70%乙醇(图1)。所有实验均采用成分近似的麦汁(15'P包含30%高麦芽糖(VHM)糖浆).并添加了ZnSO4(0.2mg/L)和硅质消泡剂(0.1mg/L)。贮藏发酵的主酵温度是18℃。
.酵母菌株
实验中所用酵母是贮藏型菌株Saccharomycescerevisiae(syn. S. pastorianus)
.酵母数
细胞数测定采用Neubauer血细胞计数器(WeberScientific Int. , Middlesex, U. K. ),其有x 40倍的光学显微镜。酵母活性测定用次甲基紫染色法。
.糖度
样品是在5, OOOrpm(5, 000 x g)离心5分钟,而后用DMA46数码密度计( PAAR Scientific Ltd.,Lon-don)测定上清液的比重。
.FAN
FAN测定用苟三酮染色法。
.氨
氨的测定是用分光光度计。校准采用的是氯化胺和蒸馏水。显色试剂采用的是3.5%酚和0.04%硝普钠的混合物。样品吸光率是用665nm透过无菌水的波长测定的。
.氨基酸
单体氨基酸的定量运用了梯度洗脱液相色谱法及荧光检测。用这种方法能完全区分所有的氨基酸。
.双乙酞及前体物
双乙酞水平分析采用气相色谱法和电子检测器(Hewlett Packanl, Crawford Scientific, Strathaven, Lanark-shire, U. K. )。将样品在60℃预热90分钟,主要是为了将前体物转化为双乙酞。未预热的样品也进行测定,有助于计算前体物浓度。前体物测定需要扣除预热样品比未预热样品增加的体积数。
结果和讨论
(图2、图3、图4)显示了主酵期FAN衰减,降糖,pH变化,发酵麦汁中的悬浮细胞数。在空白对照发酵中(见图2),发酵24小时后悬浮细胞数浓度达到最大值9x10 7个/mL。48小时内FAN浓度持续快速衰减,而后以稳定速率减少。发酵48小时后声值从5.07降至3.78,而后维持较稳定的水平直至发酵结束。麦汁糖度(SP gr)明显下降至主酵结束。糖度和FAN浓度步调一致的降低,这两种数值变化是有相关性的。
在添加了赖氨酸补充剂的发酵中(见图3),发酵开始19小时后悬浮细胞数达到最大值13 x 10 7个/mL. FAN浓度在开始的24小时内急速下降,而后达到稳定水平。而当糖度持续降低直至发酵结束时,声的变化趋势与FAN相近。将赖氨酸作为补充剂,主要是因为它是能够被酵母完全利用的氨基酸之一。
在添加了蛋氨酸补充剂的发酵中(见图4),发酵24小时后悬浮细胞浓度达到最大值7 x 10 7个/ML。主酵全过程FAN浓度连续降低,声和糖度也呈现相同的变化趋势。蛋氨酸作为补充剂主要是由于其属于B族氨基酸,能够很快地被酵母同化利用。
(图5、图6、图7)说明了氨基酸随时间的消耗。在空白对照中,A族的所有氨基酸在发酵开始的43小时内能够完全被同化(见图5)。而在添加了赖氨酸和蛋氨酸补充剂的实验中,A族的氨基酸分别在43和72小时内完全消耗(见图6和7)。在空白实验中B族氨基酸在48小时全部消耗,而在添加蛋氨酸的发酵中则是103小时完成。在添加了赖氨酸的发酵中,是43小时内被利用。在添加了赖氨酸的实验和空白对照中,C族氨基酸均在48小时消耗。在添加了蛋氨酸补充剂的实验中,此类氨基酸无法完全吸收。
如预想的,在所有的发酵过程中脯氨酸都无法被吸收利用。在添加赖氨酸的发酵中,主酵开始15小时内麦汁中的氨能够完全利用。而在空白对照及添加蛋氨酸补充剂的发酵中,氨是无法完全利用的。
含赖氨酸补充剂的主酵过程快于空白实验及添加蛋氨酸补充剂的麦汁发酵(45小时完成)。通过悬浮酵母浓度的降低可以解释说明这种影响。赖氨酸刺激酵母生长的原因暂时无法说明,但这种氨基酸已明显成为麦汁氨基酸消耗的“钥匙”。
(图8)显示了在发酵过程中VDK及其前体物浓度的变化。在发酵过程中双乙酞,2,3-戊二酮及其前体物生成的浓度,添加了赖氨酸的近乎是空白对照的两倍。另一方面就添加了蛋氨酸补充剂的样品来看,这些风味组分及其前体物质含量均低于空白实验。
结论
麦汁中添加的I-蛋氨酸补充剂抑制了酵母发酵,并将主酵时间延长到96至103小时范围。相对于空白样品,其在主酵24小时内获得的悬浮酵母数偏低。蛋氨酸的存在阻止了C族氨基酸的吸收。发酵液中双乙酸及2,3一戊二酮的含量也低于空白实验。这种偏低的现象主要是由于蛋氨酸的过量延长了主酵期,酵母生长速率的降低也很好地解释了发酵结束时VDK水平的降低。
添加L赖氨酸增强了酵母代谢活力,将主酵时间缩短到96至48小时范围。这种影响可解释为刺激作用,因为悬浮酵母数出现了大幅提高。在发酵初始43小时内氨基酸的主要成分被利用,氨也是在主酵19小时内消耗。菌体量的增加似乎也是与含补充剂的发酵液中VDK水平的提高相关联的。
这些实验只是研究酿造酵母发酵过程中麦汁含氮组分(寡肤,钱盐,及氨基酸等)作用的一部分。对类似赖氨酸作用的“标志性”氨基酸或麦汁中其它能够引起、加强酵母发酵力的含氮物质的研究正在进行中。
译自:《KMBAA TQ》2005年第2期




 加载中...
加载中...