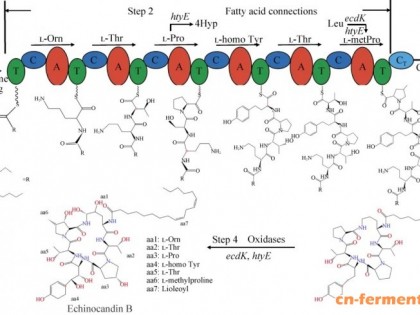
作者:田永强 赵国屏 覃重军
【摘要】 非链霉菌属的稀有放线菌类群广泛分布在自然环境中,具有重要的基础研究与应用价值。在许多稀有放线菌发现了环型与线型质粒,由质粒发展起来的遗传操作系统可以为研究稀有放线菌的生理及代谢提供强有力的工具。本文综述目前稀有放线菌中的质粒生物学的研究进展。
【关键词】 稀有放线菌 质粒
Progresses on plasmid biology of the rare actinomycetes
ABSTRACT The non?Streptomyces actinomycetes, living widespread in various habitats, are important for the basic and applied researches purposes. They harbor indigenously circular as well as linear plasmids, a useful source for developing genetic tool for studying of their unique physiology and metabolism. In this review, we summarize the current progresses on plasmid biology in the rare actinomycetes.
KEY WORDS Rare actinomycetes; Plasmid
放线菌类群是十分重要的高G+C含量的革兰阳性细菌,它们的培养形态与细菌有本质区别,大多数产生菌丝结构,包括培养基中的基质菌丝和培养基表面的气生菌丝及孢子丝。迄今已发表145个属,约2000个种,其中链霉菌属中约500个种。它们产生了大约10000种抗生素和生理活性物质,在医药、工业和农业方面具有重要的应用价值[1]。非链霉菌属的稀有放线菌广泛分布于各种土壤环境中,常具有独特的形态及生理特点。稀有放线菌也象链霉菌一样能够产生丰富的次生代谢产物,如拟无枝菌酸菌产生的利福霉素(rifamycin)及万古霉素(vancomycin)[2],游动放线菌产生的替考拉宁(teicoplanin)[3]等在临床上是极为重要的抗生素。此外,许多稀有放线菌属在农业、环保和医学方面具有重要的意义,如可以进行生物固氮的弗兰克菌,具有降解有毒害化合物功能的红球菌,导致人类麻风病和结核病的分枝杆菌等[4]。过去几十年里,放线菌的遗传学研究主要集中在链霉菌属,对放线菌其它类群的遗传学研究相对较少,尤其是对它们的遗传操作体系则更少。对稀有放线菌属的生理或病理的研究迫切需要分子操作工具,所以发展稀有放线菌的遗传操作就有非常现实的意义。本文综述非链霉菌属的放线菌质粒生物学及其遗传操作体系的研究进展。
1 稀有放线菌中的质粒研究现状
1.1 分枝杆菌质粒
分枝杆菌作为重要的病原菌成为许多实验室的研究对象。自1979年Crawford等[5]第一次从分枝杆菌Mycobacterium avium发现环型质粒存在后[5],陆续在M.avium、 M.intracellulare和M.scrofulaceum及快速生长菌M.fortuitum、M.peregrinum、M.abscessus和M.chelonae中报道了许多从13kb到150kb大小不等的质粒[6,7],如从M.avium中分离得到15.3kb的质粒pRL7[8],从M.fortuitum中分离到pAL5000[9]和单拷贝的pMF1[10],从M.scrofulaceum中得到了pMSC262[11]。M.scrofulaceum和M.avium对水银及铜的耐受性,以及质粒介导的限制性修饰均与质粒的存在有关[12,13]。Beggs等发现pRL7的序列与pMSC262具有高度同源,它们可能属于同一不相容群[8],而pMSC262与pAL5000具有相容性[11]。几个分枝杆菌的环型质粒的复制起始区被确定,并用来构建能在大肠埃希菌和分枝杆菌中穿梭的载体[8,9,11]。然而这些载体的功能仍然有其局限性,比如它不能在快速生长的常用宿主菌M.smegmatis mc#155中复制[14]。只有pAL5000衍生的质粒在快速生长与慢速生长的分枝杆菌菌株中都具有复制活性。
除环型质粒外,Picardeau等报道了在M.xenopi、M.branderi、M.celatum type 2和M.celatum type 3等菌株中发现14个线型质粒,大小从20kb到320kb,通过Southern blotting分析可以将它们分为不同的类群,同类群的质粒是不相容的[15]。Picardeau等对M.celatum来源的25kb线型质粒pCLP进行了分析,发现它具有与其它放线菌线型质粒相同的端粒结构[16]。同时它们对质粒pCLP的复制区功能进行了测定,复制区有两个ORF,分别编码复制和分配蛋白,其间有一小段与分枝杆菌环型质粒复制起始区相似的序列,推测其为线型质粒的复制起始区。用该复制区构建的载体pCL4D与M.fortuitum来源的环型质粒pAL5000和pJAZ38具有相容性。
1.2 棒状杆菌质粒
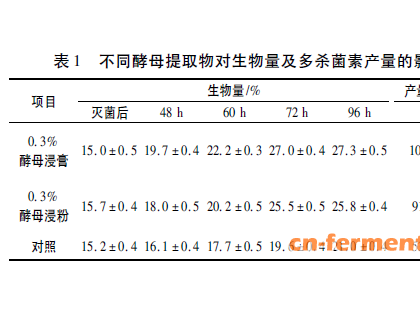
棒状杆菌由非致病的土壤类群(Corynebacterium glutamicum,C.callunae,Brevibacterium lactofermentum)、动物致病类群(C.diphtheriae,C.xerosis)和植物致病类群(C.nebraskense,C.michiganense)组成。非致病的土壤棒状杆菌在工业上主要用于生产氨基酸,另外的用途包括降解石腊(paraffin)生产去污剂及固醇转化等。从20世纪70年代末期开始,许多实验室进行了大量的筛选工作找到了许多大小分布在1.4kb到69kb之间的质粒[17~20]。通常这些菌株的大质粒上携带有抗生素耐受性基因(表1)。
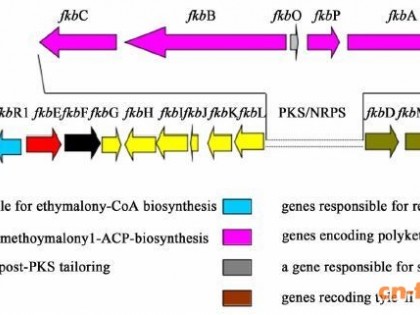
Tauch等从氨基酸产生菌C.glutamicum中分离到24个2.4~95kb大小不等的质粒,根据质粒的复制起始方式与复制起始蛋白的序列同源性可以将这些质粒划分为4个类群[21]。 最近几年来, 对质粒的研究工 表1 棒状杆菌中携有耐受的质粒作主要集中在对pBL1、pSR1、pGA1和pCG100等质粒的最小复制子及复制机制的研究[22]。通过对这些质粒的复制区结构的解析,了解它们不同的复制方式,如pBL1和pSR1采用滚环复制方式,而pRBL1则采用theta方式复制(表2)。
Katsumata[23]和Martin[24]分别用不同的质粒发展了棒状杆菌的克隆载体及转化系统。主要的克隆载体是由pBL1和pSR1复制子构建而来[24,25]。大肠埃希菌中的启动子序列与棒状杆菌的具有同源性,因此,大肠埃希菌中的lac,trp、PL操纵子和PR阻遏子可以在棒状杆菌中表达,这就为构建棒状杆菌的表达载体提供了便利[26,27]。
1.3 诺卡菌质粒
诺卡菌作为好气的放线菌是引起人体感染的病原菌。但是对它的质粒生物学性质研究并不多。Kirby等[28]于1985年第一次从诺卡菌Nocardia corallina中分离到2.7kb的环型质粒pKU100。之后,诺卡菌质粒的工作停滞不前,许多诺卡菌的基因都可在链霉菌中进行表达[29]。Madon等利用地中海拟无枝菌酸菌(Amcolatopsis mediterranei)质粒pMAE100发展出能在诺卡菌中复制的载体pMAE123[30]。最近Liu等[31]在N.italica中发现环型质粒pNI100,并克隆了该质粒的复制起始区, 该区域包含一个编码396个氨基酸
表2 棒状杆菌环型质粒复制模式
类群质 粒 复制方式与Rep蛋白同源的质粒ⅠpBL1滚环复制 pIJ101 (Streptomyces lividans) pSB24 (S.cyanogenus) pJV1 (S.phaerochromogenes)ⅡpSR1,pNG2滚环复制pGA1ⅢpRBL1Theta复制 pAL5000 (Mycobacterium fortuitum) pCo1E5?099 (Shigella sonnet) pJD1 (Neisseria gonorrhoene)
的蛋白,与链霉菌质粒pSG5的Rep蛋白有3段同源区域,推测它可能具有与pSG5相似的滚环复制机制。该质粒可以通过转化变铅青链霉菌(S.lividans)ZX7而产生麻点(pock)现象。根据pNI100复制子发展了能在大肠埃希菌与诺卡菌穿梭的质粒pNI105,这为诺卡菌的遗传操作提供了便利。
Kalkus等[32]在氢自养型N.opaca中发现了多个线型和环型质粒,如菌株MR11含有环型质粒pHG31?a和pHG31?b及线型质粒pHG201(270kb)、pHG202(400kb)、pHG203(420kb),菌株MR22中环型质粒pHG33和pHG204(180kb)、pHG205(280kb)和pHG206(510kb)。用氢化酶与二磷酸核酮糖羧化酶基因作为探针进行交分析,在接合转移质粒pHG201与pHG205中均发现阳性信号,证实它们与氢的代谢有关。
1.4 红球菌质粒
在过去20年里,红球菌作为与环境相关的微生物受到越来越多的关注。它们可以降解许多环境污染物,如酚类物质、杀虫剂、除草剂、丙稀酰胺、苯胺、卤化烷烃等有害物质[33~37]。目前从红球菌中分离到许多环型质粒,这些质粒按复制子的类型可分为两种,其一,以theta方式复制的质粒,如pRE2895。该质粒编码RepA和RepB与ColE2型质粒的复制蛋白具有相同的特点,故推测其具有theta方式的复制机制。目前利用pRE2895的repAB构建了表达质粒pTip[38]。其二,以滚环方式复制的质粒,如R.erythropolis DSM8424的pRE8424。它有一个单链起始区和一个双链起始区,这是典型的滚环复制单元。对单链起始区的突变分析发现单链中间体在体内积累,这是因为单链不能及时转化成双链引起的。以pRE8424复制区构建的表达质粒能与以pRE2895为基础的质粒同时存在细菌体内,表明它们属于不同的不相容群[39]。
对于R.aetherovorans I24(7Mb)和R.erythropolis PR4 (7Mb)的基因组研究表明红球菌体内存在许多大的线型质粒,许多与环境污染物降解有关的基因就位于线型质粒上[40]。如多氯联苯(PCB)的降解菌R.erythropolis RHA1包含三个线型质粒:pRHL1(1100kb)、pRHL2(450kb)和pRHL3(330kb),PCB的降解基因bph分散在染色体与质粒pRHL1和pRHL2上[41]。在R.erythropolis BD2线型质粒pBD2(210kb)上还发现异丙基苯的降解基因ipb和具有降解联苯降解功能的etbD1类似基因[42]。此外,催化有机硫化物脱硫的催化途径位于红球菌IGTS8的150kb的线型质粒上[43],红球菌DK17具有两个线型质粒pDK1(380kb)和pDK2(330kb),后者含有烷基苯代谢的单加氧酶与meta?环裂解的双加氧酶基因[44]。
1.5 弗兰克菌质粒
在弗兰克菌中已发现多个环型质粒,大小从7kb到190kb[45~47]。目前已完成了来自菌株ArI3的质粒pFQ31[48]和菌株CpI1的pFQ11[49]以及pFQ12[50]的测序工作。22.4kb的pFQ12有超过50个开放阅读框(ORF),9个有转录活性[50]。pFQ31有17个ORF[48],pFQ11有4个[49],pFQ11与pFQ31有大约99.7%的同源性。在3个质粒上都发现了质粒分配蛋白基因,如pFQ31的parFA、pFQ11的ORF3编码类似假单胞菌Pseudomonas alcaligenes ParA的蛋白,pFQ12含有一个蛋白类似芽孢杆菌B.subtilis的SpoJ。这些基因的编码产物使质粒能在子细胞中稳定存在。同时还发现与类似链霉菌(S.ambofaciens)中接合转移与整合性质粒pSAM2 KorSA的蛋白。根据ptrFA和repFA之间的DNA结构域推测质粒pFQ31的复制起始区位于其间,该区域序列具有类似重复子(iteron)的序列,这些结构通常在以theta方式复制质粒的复制区中存在,repFA编码复制起始蛋白Rep。pFQ12有两个开放阅读框(ORF3与ORF6)可能与质粒的转移有关。
1.6 小单孢菌质粒
小单孢菌(Micromonospora)是多种重要抗生素的产生菌[51],其遗传操作主要依赖于噬菌体构建的载体[52]。Parag等[53]首先报道从西索米星(sisomicin)产生菌M.inyioensis分离到环型质粒。随后Oshida[54]报道从M.zionensis NRRL5466(西索米星产生菌)中找到高拷贝质粒pMZ1(9.9kb),从M.rosaria NRRL3718(罗沙米星产生菌)发现53.5kb质粒pMR1和11.0kb小质粒pMR2。Hasegawa等[55]利用小单孢菌的质粒复制子构建了一系列载体pMO116、pMO126、pMO133、pMO136和pMO217。Vukov等[56]利用pMZ2的复制区构建了接合转移载体pMZS25和pMZS34。
1.7 拟无枝菌酸菌质粒
拟无枝菌酸菌(Amycolatopsis)最早被划分为链霉菌属,之后归为诺卡菌属,到20世纪80年代中期,Lechecalier等[57]根据细胞壁组成缺乏以及对诺卡菌和红球菌的噬菌体不敏感等特征将其归为拟无枝菌酸菌属。拟无枝菌酸菌能产生利福霉素、万古霉素和balhimycin等在临床上重要的抗生素。拟无枝菌酸菌的遗传操作系统还不够成熟,造成对其遗传学的研究进展缓慢。许多来源于链霉菌的质粒,如pIJ702和来源于诺卡菌的质粒pCY104都不能在该菌中稳定的复制和表达。Moretti等从A.mediterranei中检测到23.7kb的质粒pMEA100,但由于容易整合到染色体上,不易分离,以及拷贝数太低,很难改造为合适的载体。Oh等[58]从糖肽类抗生素万古霉素的产生菌A.orientalis中分离到33.5kb的质粒,但是该质粒不能稳定地遗传。Lal等[59]从一株拟无枝菌酸菌中分离到一个约29.6kb的质粒pA387。其衍生载体pRL1转化到N.lactamdurans中后,自发地丢失一个片段得到质粒pULVK2。
1.8 其它稀有放线菌的质粒
马杜拉放线菌产生多种抗生素[60],但是其遗传操作体系的发展却相对落后,原因在于从马杜拉菌中发现的质粒相对较少。只有Dairi等[61]分离得到4个质粒(pAE042、pTPA0016、pTPA0019和pTPA0123),并利用pTPA0123的复制区构建了能在Actinoma?dura verrucosospora中稳定复制的克隆载体。Fare等[62]从链孢囊菌Streptosporangium brasiliense中分离到环型质粒pSgB?1,从S.viridognriseum中分离到质粒pSgV?1。Polo等[63]从延伸因子Tu抑制剂的产生菌玫瑰游动双孢菌(Planobispora rosea)中分离得到两个线型质粒pPR1(27.5kb)和pPR1(16kb)。两个质粒具有相似的序列,但它们的端粒结构有一定的差异,pPR2端粒区域的650个碱基具有类似于链霉菌端粒的倒转重复序列。
2 前景展望
稀有放线菌产生的抗生素种类占所有放线菌产抗生素的三分之一[62],随着对稀有放线菌环型与线型质粒被发现,越来越多的克隆或表达载体被构建,可以为深入研究稀有放线菌的遗传特征提供更多的帮助,也为有效利用微生物资源提供了有力的工具。
【参考文献】
[1] 刘志恒,姜成林. 放线菌现代生物学与生物技术[M]. 北京:科学出版社,2004
[2] Yao Y, Zhang W, Jiao R, et al. Efficient isolation of total RNA from antibiotic?producing bacterium Amycolatopsis mediterranei [J]. J Microbiol Methods,2002,51(2):191
[3] Sosio M, Bianchi A, Bossi E, et al. Teicoplanin biosynthesis genes in Actinoplanes teichomyceticus [J]. Antonie Van Leeuwenhoek,2000,78(3~4):379
[4] Cosma C L, Sherman D R, Ramakrishnan L. The secret lives of the pathogenic mycobacteria [J]. Annu Rev Microbiol,2003,57:641
[5] Crawford J T, Bates J H. Isolation of plasmids from mycobacteria [J]. Infect Immun,1979,24:979
[6] Jensen A G, Bennedsen J, Rosdahl V T. Plasmid profiles of Mycobacterium avium/intracellulare isolated from patients with AIDS or cervical lymphadenitis and from environmental samples [J]. Scand J Infect Dis,1989,21:645
[7] Labidi A, Dauguet C, Goh K S, et al. Plasmid profiles of Mycobacterium fortuitum complex isolates [J]. Curr Microbiol,1984,11:235
[8] Beggs M L, Crawford J T, Eisenach K D. Isolation and sequencing of the replication region of Mycobacterium avium plasmid pLR7 [J]. J Bacteriol,1995,177:4836
[9] Ranes M G, Rauzier J, Lagranderie M, et al. Functional analysis of pAL5000, a plasmid from Mycobacterium fortuitum: construction of a "mini" mycobacterium?Escherichia coli shuttle vector [J]. J Bacteriol,1990,172:2793
[10] Bachrach G, Colston M J, Bercovier H, et al. A new single?copy mycobacterial plasmid, pMF1, from Mycobacterium fortuitum which is compatible with the pAL5000 replicon [J]. Microbiology,2000,146:297
[11] Qin M, Taniguchi H, Mizuguchi Y. Analysis of the replication region of a mycobacterial plasmid, pMSC262 [J]. J Bacteriol,1994,176:419
[12] Meissner P S, Falkinham III J O. Plasmid?encoded Mercuric reductase in Mycobacterium scrofulaceum [J]. J Bacteriol,1984,157:669
[13] Erardi F X, Failla M L, Falkinham III J O. Plasmid?encoded copper resistance and precipitation by Mycobacterium scrofulaceum [J]. Appl Environ Microbiol,1987,53:1951
[14] Snapper S B, Melton R E, Mustafa S, et al. Isolation and characterization of efficient plasmid transformation mutants of Mycobacterium smegmatis [J]. Mol Microbiol,1990,4:1911
[15] Picardeau M, Vincent V. Characterization of large linear plasmids in mycobacteria [J]. J Bacteriol,1997,179:2753
[16] Picardeau M, Vincent V. Mycobacterial linear plasmids have an invertron?like structure related to other linear replicons in actinomycetes [J]. Microbiology,1998,144:1981
[17] Kaneko H, Tanaka T, Sakaguchi K. Isolation and characterization of a plasmid from Brevibacterium lactofermentum [J]. Agric Biol Chem,1979,43:867
[18] Gross D C, Vidaver A K, Keralis M B. Indigenous plasmids from phytopathogenic Corynebacterium sp [J]. J Gen Microbiol,1979,115:479
[19] Kono M, Sasatsu M, Aoki T. R plasmid in Corynebacterium xerosis strains [J]. Antimicrob Agents Chemother,1983,23:506
[20] Martin J F, Santamaria R, Sandoval H, et al. Cloning systems in amino acid producing corynebacteria [J]. Bio Technology,1987,5:137
[21] Tauch A, Puhler A, Kalinowski J, et al. Plasmids in Corynebacterium glutamicum and their molecular classification by comparative genomics [J]. J Biotechnol,2003,104:27
[22] Santamaria R, Gil J A, Mesas J M, et al. Characterization of an endogenous plasmid and development of cloning vectors and a transformation system in Brevibacterium lactofermentum [J]. J Gen Microbiol,1984,130:2237
[23] Katsumata R, Ozaki A, Kon T, et al. Protoplast transformation of glutamate producing bacteria with DNA [J]. J Bacteriol,1984,159:306
[24] Santamaria R, Gil J A, Mesas J M. Characterization of an endogenous plasmid and development of cloning vectors and a transformation system in Brevibacterium lactofermentum [J]. J Gen Microbiol,1984,130:2237
[25] Takagi H, Morinaga Y, Miwa K, et al. Versatile cloning vectors constructed with genes indigenous to glutamic acid producer Brevibacterium lactofermentum [J]. Agric Biol Chem,1986,50:2597
[26] Liebl W, Sinskey A J, Schleifer K. Expression, secretion and processing of staphylococcal nuclease by Coryne?bacterium glutamicum [J]. J Bacteriol,1992,171:1854
[27] Billman?Jacobe H, Hodgson A L M, Lightowlers M, et al. Expression of ovine gamma interferon in Escherichia coli and Corynebacterium glutamicum [J]. Appl Environ Microbiol,1994,60:1641
[28] Kirby R, Usdin K. The isolation and restriction mapping of a miniplasmid from the Actinomycete Nocardia coralline [J]. FEMS Microbiol Lett,1985,27:57
[29] Enguita F J, Fuente J L de la, Martín J F, et al. An inducible expression system of histidine?tagged proteins in Streptomyces lividans for one?step purification by Ni2+ affinity chromatography [J]. FEMS Microbiol Lett,1996,137:135
[30] Madon J, Hutter R. Transformation system for Amycolatopsis (Nocardia) mediterranei: direct transformation of mycelium with plasmid DNA [J]. J Bacteriol,1991,173:6325
[31] Liu Y T, Su C M, Lee C H, et al. Cloning and characterization of the replicon of the Nocardia italica plasmid, pNI100 [J]. Plasmid,2000,43:223
[32] Kalkus J, Reh M, Schlegel H G. Hydrogen autotrophy of Nocardia opaca strains is encoded by linear megaplasmids [J]. J Gen Microbiol,1990,136:1145
[33] Straube G, Hensel J, Niedan C, et al. Kinetic studies of phenol degradation by Rhodococcus sp. P1. 1. Batch cultivation. Antonie van Leeuwenhoek [J]. J Microbiol Serol,1990,57:29
[34] Ferguson J A, Korte F. Epoxidation of aldrin to exo?dieldrin by soil bacteria [J]. Appl Environ Microbiol,1981,34:7
[35] Behki R, Topp E, Dick W, et al. Metabolism of the herbicide Atrazine by Rhodococcus strains [J]. Appl Environ Microbiol,1993,59:1955
[36] Kobayashi M, Nagasawa T, Yamada H. Enzymatic synthesis of acrylamide: a success story not yet over [J]. Trends Biotechnol,1992,10:402
[37] Sallis P J, S. Armfield J, Bull A, et al. Isolation and characterization of a haloalkane halidohydrolase from Rhodococcus erythropolis Y2 [J]. J Gen Microbiol,1990,136:115
[38] Nakashima N, Tamura T. A novel system for expressing recombinant proteins over a wide temperature range from 4 to 35 degrees C [J]. Biotechnol Bioeng,2004,86:136
[39] Nakashima N, Tamura T. Isolation and characterization of a rolling?circle?type plasmid from Rhodococcus erythropolis and application of the plasmid to multiple?recombinant?protein expression [J]. Appl Environ Microbiol,2004,70:5557
[40] van der Geize R, Dijkhuizen L. Harnessing the catabolic diversity of rhodococci for environmental and biotechnological applications [J]. Curr Opin Microbiol,2004,7:255
[41] Shimizu S, Kobayashi H, Masai E, et al. Characterization of the 450?kb linear plasmid in a polychlorinated biphenyl degrader, Rhodococcus sp. strain RHA1 [J]. Appl Environ Microbiol,2001,67:2021
[42] Stecker C, Johann A, Herzberg C, et al. Complete nucleotide sequence and genetic organization of the 210?kilobase linear plasmid of Rhodococcus erythropolis BD2 [J]. J Bacteriol,2003,185:5269
[43] Denis?Larose C, Labbe D, Bergeron H, et al. Conservation of plasmid?encodeddibenzothiophene desulfurization genes in several rhodococci [J]. Appl EnvironMicrobiol,1997,63:2915
[44] Kim D, Kim Y S, Kim S K, et al. Monocyclic aromatic hydrocarbon degradation by Rhodococcus sp. strain DK17 [J]. Appl Environ Microbiol,2002,68:3270
[45] Dobritsa S V. Extrachromosomal circular DNA in endosymbiont vesicles from Alnus glutinosa root nodules [J]. FEMS Microbiol Lett,1982,15:87
[46] Normand P, Simonet P, Butour J L, et al. Plasmids in Frankia sp. [J]. J Bacteriol,1983,155:32
[47] Simonet P, Normand P, Moiroud A, et al. Restriction enzyme digestion patterns of Frankia plasmids [J]. Plant Soil,1985,87:49
[48] Lavire C, Louis D, Perriere G, et al. Analysis of pFQ31, a 8551?bp cryptic plasmid from the symbiotic nitrogen?fixing actinomycete Frankia [J]. FEMS Microbiol Lett,2001,197:111
[49] Xu X, Kong R, de Bruijn F J, et al. Sequence and characterization of plasmid pFQ11 and glutmine synthetase gene glnA from Frankia spp. [A]. 11th International Meeting on Frankia and Actinorhizal plants. Carry?le?Rouet, France,2001
[50] John T R, Rice J M, Johnson J D. Analysis of pFQ12; a 22.4kb Frankia plasmid [J]. Can J Microbiol,2001,47:1
[51] Wagman G H, Weinstein M J. Antibiotic from Micro?monospora [J]. Annu Rev Microbiol,1980,34:537
[52] Li X, Zhou X, Deng Z. Isolation and characterization of Micromonospora phage PhiHAU8 and development into a phasmid [J]. Appl Environ Microbiol,2004,70:3893
[53] Parag Y, Goedeke M E. A plasmid of the sisomicin producer Micromonospora inyoensis [J]. J Antibiot (Tokyo),1984,37:1082
[54] Oshida T, Takeda K, Yamaguchi T, et al. Isolation and characterization of plasmids from Micromonospora zionensis and Micromonospora rosaria [J]. Plasmid,1986,16:74
[55] Hasegawa M, Dairi T, Ohta T, et al. A novel highly efficient gene?cloning system for Micromonospora strains [J]. J Bacteriol,1991,173:7004
[56] Vukov N, Vasiljevic B. Analysis of plasmid pMZ1 from Micromonospora zionensis [J]. FEMS Microbiol Lett,1998,162:317
[57] Lechevalier M P, Prauser H, Labda D P, et al. Two new genera of nocardioforma actinomycetes: Amycolata gen nov. and Amycolatopsis gen nov. [J]. Int J Syst Bac?teriol,1986,36:29
[58] Oh Y K, Fare L R, Taylor D P, et al. A cryptic plasmid from Nocardia orientalis NRRL 2452, a vancomycin producer [J]. J Antibiot (Tokyo),1986,39:694
[59] Lal R, Khanna M, Kaur H, et al. Rifamycins: strain improvement program [J]. Crit Rev Microbiol,1995,21:19
[60] Miyadoh S. Research on antibiotic screening in Japan over the Last decade: a producing microorganism approach [J]. Actinomycetologica,1993,7:100
[61] Dairi T, Hamano Y, Furumai T, et al. Development of a self?cloning system for Actinomadura verrucosospora and identification of polyketide synthase genes essential for production of the angucyclic antibiotic pradimicin [J]. Appl Environ Microbiol,1999,65:2703
[62] Fare L R, Taylor D P, Toth M J, et al. Physical characterization of plasmids isolated from Streptosporangium [J]. Plasmid,1983,9:240
[63] Polo S, Guerini O, Sosio M, et al. Identification of two Linear plasmids in the actinomycete Planobispora rosea [J]. Microbiology,1998,144:2819