


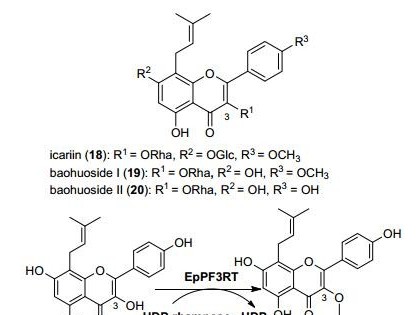
 13720223167
13720223167



与普通微生物一样,基因重组菌的发酵生产水平不但和菌种的遗传特性有关,也和发酵工艺控制以及发酵罐的性能有着密切的关系。合理的发酵工艺控制可以给微生物提供良好的环境,提高细胞水平的调控,提高目标产品的生产水平。蛋白酶是工业酶种中用的最多的一种酶,约占酶总量的60%,而碱性蛋白酶又占了蛋白酶的市场销售额的50%以上,它在洗涤剂、食品工业和皮革制造中发挥巨大作用,目前,国内外市场上对碱性蛋白酶的需求量增长很快。自从Jacobs通过建立基因文库克隆了第一条碱性蛋白酶基因以来,利用基因工程的手段和蛋白质工程技术获得高产碱性蛋白酶的工程菌已经是当前蛋白酶研究中的一个热点,这给蛋白酶的进一步应用研究提供了广阔的前景[1]。我们在前期工作中利用地衣芽孢杆菌为宿主获得一株碱性蛋白酶的整合型工程菌BA071,使其酶产量在基础培养基下提高了85%。国外生产菌株目前均为基因工程菌株,发酵单位产量在25000~30000u/mL之间[2]。我国的生产菌株的发酵水平有文献报道的最高不超过18000u/mL[3]。目前国内外的相关文献中,多是以对传统诱变筛选获得的生产碱性蛋白酶的芽孢杆菌进行发酵条件的研究,对重组碱性蛋白酶工程菌进行发酵条件的的研究还很少。本实验即是在5L发酵罐上对所构建的碱性蛋白酶整合型基因工程菌产酶发酵条件进行了研究,探讨了相关工艺参数的变化。为了给探索重组酶的性质及其稳定性奠定基础,我们利用快速蛋白液相层析,即FPLC(FastProteinLiquidChromatography)技术,建立了快速高效纯化碱性蛋白酶的方案,并对其重组酶的酶学性质进行了研究。
1 材料与方法
1.1 菌株
整合型基因工程菌BL071为前期工作构建[4]。
1.2 培养基
LB培养基(1000mL):酵母粉5g,蛋白胨10g,氯化钠10g,pH7.2~7.4;
干酪素固体培养基(1000mL):干酪素8g,酵母粉2g,KH2PO414g,K2HPO46g,(NH4)2SO42g,琼脂12g,pH7.2~7.4;
基础产酶培养基:按参考文献[5],自然pH;
发酵培养基(1000mL):黄豆饼粉50g,玉米粉50g,麸皮25g,KH2PO40.3g,Na2HPO40.4g,Na2CO31g;自然pH,在36℃下通风培养,前期通风1:0.15,后期通风1:0.2,搅拌40h左右。
1.3 主要仪器和试剂
全自动发酵罐KF 5L(韩国发酵机株式会社产品);快速蛋白液相层析系FPLC,垂直板状电泳系统(Bio Rad公司产品);DEAESephadex A 50、CM Seph adex C 50、Sephadex G 75均为瑞典Pharmacia公司产品,其他试剂均为国产分析纯。
1.4 发酵罐发酵条件
发酵罐有效容积5L,投料体积2L;培养温度36℃,pH值和DO值采用pH电极和溶氧电极在线检测。
1.5 细胞生长曲线
测定采用血球计数法测定取平均值。
1.6 还原糖测定
培养液离心、过滤去上清,用生物传感器检测。硫酸铵梯度盐析、透析和蛋白质浓缩方法见文献[6]。
1.7 CM Sephadex C 50柱层析将处理好的凝胶装柱后,用0.02mol/LpH6.0的PBS缓冲液平衡,上样后用同样的PBS缓冲液(流经A泵)及含1mol/LNaCl的0.02mol/LpH6.0的PBS缓冲液(流经B泵)在FPLC系统中自动混匀进行离子梯度洗脱,分步收集器速度为4mL/min,收集活性峰洗脱液。
1.8 Sephadex G 75柱层析
将CM Sephadex C 50层析所收集活性峰洗脱液2mL上样,用含0.15mol/LNaCl的0.02mol/LpH6.0的PBS缓冲液在FPLC系统控制下洗脱,分步收集器速度为2mL/min收集活性峰,浓缩酶液,至-4℃保存。
1.9 蛋白浓度测定Bradford法[7],以牛血清白蛋白作为标准。
1.10 SDS PAGE分析
SDS PAGE方法参照文献[8],采用12%的分离胶和4%的浓缩胶进行。
1.11 酶活测定方法
按部颁行业标准QB 1803[5]进行酶活定义:1mL液体酶,在40℃和pH10.5的条件下,1min水解1μg酪素产生1μg酪氨酸为1个活力单位,以u/mL表示。
2 结果与讨论
2.1 不同转速对重组菌产酶的影响
地衣芽孢杆菌产碱性蛋白酶是耗氧发酵过程,可以改变搅拌转速来调节通气量。搅拌转速的增加,在一定范围内可以显著提高溶氧,搅拌转速超过一定值后就不在成为提高溶氧的限制因素。剧烈的搅拌除了消耗大量功率外,还会使发酵液产生大量泡沫,搅拌叶的剪切作用也会造成细胞的损伤,使菌体提前自,产酶量降低。因此,对产酶而言,存在一个最佳搅拌转速的问题。搅拌对泡沫的影响很大。一般发酵罐接种后加大风量和搅拌,会使泡沫多到无法控制。一般发酵前期风量不需要很大,此时菌体呼吸虽然旺盛,但菌体数量尚少,总的耗氧量不高,应先开小风量逐步加大,待菌体浓度达到一定程度以后,再加大风量,同时用消泡剂消除泡沫。通过预备实验确定本实验中采用前期通风1:0.15,后期通风1:0.2,结果显示(图1),在500r/min搅拌转速下发酵水平最高,为24480u/mL;700r/min搅拌转速下发酵水平次之,300r/min搅拌转速下发酵水平最低。而原出发菌株在500r/min搅拌转速下,发酵水平为9040u/mL;重组菌的发酵水平比出发菌株提高了170.8%。

2.2 5L发酵罐培养条件下生长曲线的测定
碱性蛋白酶是在菌体生长对数后期分泌表达,并随着芽孢的形成而快速消失。在500r/min下,测定发酵过程中菌体的生长情况。从图2可知,菌体在24~32h内生长迅速,以后细胞略有增长,36h左右处于稳定期。在充分溶氧的条件下,菌体密度最高可达到87亿个/mL(因为培养基粘度很大,杂质较多,无法用OD值法以及细胞干重法检测,所以采用细胞计数法)。与图1中结果比较可知,菌体的生长和酶活的变化趋势基本一致,菌体生长到40h左右达到最大值,酶活最高值在40h左右达到最高,因此可以确定发酵终点为40h。
2.3 发酵过程中还原糖及pH的变化
从图3中可以看出,培养基中的还原糖随着细胞需求量的增大而逐渐下降。在20~30h细胞生长速度加快,细胞对氧的需求加大,所以此时较高搅拌速度可以改善溶氧,否则发酵液中往往会形成溶氧低谷,甚至DO值掉零,从而大大影响细胞的生长,难以提高菌体的密度。
2.4 重组酶的制备及其纯化
重组酶的纯化的过程如表1所示。重组酶纯度提高了76.2倍。
2.5 SDS PAGE分析收集Sephadex G 75层析所得的活性峰酶液浓缩,取10μL浓缩液上样。结果显示得到单一电泳条带,分子量为28kD(图4)。
2.6 重组酶的最适pH
分别采用H3PO4 NaH2PO4(pH4~5)、NaH2PO4 Na2HPO4(pH6~8)Na2B4O7.NaOH(pH9~14)组成的缓冲体系[9],用0.1mol/L的不同pH的缓冲液配制成1%浓度的酶液,以酪蛋白为底物,在40℃反应10min。测定纯酶在一系列pH下的活力(图5)。结果表明,酶的最适pH在11左右。
2.7 重组酶的最适温度
用不同pH11缓冲液配制的酶液,以酪蛋白为底物在不同温度中反应10min,测定纯酶在不同温度下的催化活力。结果表明(图6),该酶在55℃~65℃之间活力最高,而酶在40℃时的活力仅为60℃时的37.5%左右。
2.8 pH对重组酶的稳定性的影响
用不同pH的缓冲液配制的酶液,40℃处理60min,再用pH11的缓冲液将其稀释一定浓度,并且使pH值均达到同一测定值,测剩余酶活力。结果表明(图7),酶在pH6.5~12.0之间均保持80%以上的活力,说明该酶具有较强的耐碱性。
2.9 重组酶的热稳定性研究
将纯酶用pH11的缓冲液稀释后中,经过不同温度保温,按一定时间间隔在40℃下测定其剩余活力。结果见图8,55℃保温2h,酶活力依然保持在80%以上,随着温度的升高,酶的热稳定性下降,60℃时,酶的半衰期为2h。65℃保温20min,剩余活力为33.7%。说明重组酶具有较好的热稳定性。
2.10 金属离子对重组酶活力的影响
用经过稀释的酶液将金属离子配制成1mmol/L的溶液,总体积2.5mL。40℃保温10min后测酶活。每组均设对照管(无金属离子)。
结果表明(表2),Ca2+、Mg2+对酶稍有激活作用,而Hg2+、Ag+对酶有明显抑制作用。Nakamura[10]等指出,Ca2+对许多酶都有稳定保护作用,尤其对耐热酶的热稳定性有很强的保护作用,因为Ca2+能帮助酶的三级结构更加稳定。Mg2+是否有这样的作用目前的研究尚不明确。
2.11 化学试剂对重组酶的影响
将不同的化学试剂与酶液混合,使化学试剂的终浓度为1×10-3mol/L,40℃保温10min后测酶活。每组均设对照管。
结果表明(表3),酶的活性被苯甲基磺酰氟(PMSF)和二异丙基氟磷酸(DFP)强烈抑制,因为其活性中心为丝氨酸残基,化学修饰后引入的基团阻碍了底物与活性中心的结合,或改变了活性中心附近的电荷性质从而影响酶的活性,这方面的研究目前尚无明确的结论。乙二胺四乙酸(EDTA)对酶稍有激活作用,而表面活性剂SDS和Urea对酶活的影响不大。
基因重组菌在构建时候,通常都是采用摇瓶培养的方法比较其生产能力,这样得到的培养工艺一般不能直接应用于发酵罐生产,这是因为在摇瓶中难以进行深入的发酵特性研究,而且摇瓶的操作方式和提供的培养环境与发酵罐有很大的差别,因此对基因重组菌进行发酵特性研究,对发酵过程加以优化,同时掌握基因重组菌发酵放大的特点,可以充分发挥基因重组菌的生产能力,保证产业化的顺利进行。为了给重组菌提供必需的营养和适宜的环境,既要考虑发酵过程中菌体的生长又要考虑外源基因的表达。一般进行基因重组菌发酵都希望得到较高的菌体密度,这样无论基因表达产物是分泌的还是积累在胞内,较高的菌体密度下产物的生产速率可达到较高的水平。
本实验在5L发酵罐中对重组碱性蛋白酶工程菌株BP071高产碱性蛋白酶的条件进行了研究,在36℃,500r/min条件下,BP071在发酵培养基中培养40h后,发酵液中碱性蛋白酶为24480u/mL。在相同发酵条件下,重组菌的发酵水平比出发菌株提高了170.8%。发酵水平基本达到国外的基因工程菌的水平。
在工业微生物的基因工程研究中,利用芽孢杆菌作为的表达宿主进行外源蛋白的分泌表达很有意义,目的蛋白能以活性结构生成而不发生聚集,避免了复性的困难,产物与大量的细胞蛋白分离,给下游工程的研究带来了便利。碱性蛋白酶为胞外酶,重组酶大量存在于发酵液中,经过梯度盐析、透析、DEAE纤维素柱脱色,及CM Sephadex C 50和Sepha dex G 75层析后获得电泳纯,纯度提高了76.2倍。由于碱性蛋白酶的pI一般在7.0~8.0之间,所以我们选用阳离子交换柱CM Sephadex C 50,并选用比碱性蛋白酶pI低不少于一个单位的pH为6.0的0.02mol/LPBS缓冲液进行洗脱,获得较好的效果。
重组酶基本酶学性质研究表明,重组酶具有较高的比活,耐碱性耐热性较好,受表面活性剂的影响较小,非常适合做洗涤剂中的添加剂。该酶的基因序列与基础研究较为深入的SubtilisinE同源性很高,也为丝氨酸碱性蛋白酶,二者在结构上和催化机制上存在相似性。有学者对SubtilisinE基因序列用定点突变等方法对蛋白质活性中心的部位或临近部位进行改造,试图获得既有高比活又能耐强氧化的新型碱性蛋白酶[11],这一方向的工作目前正成为基础研究的热点,也为以地衣芽孢杆菌为宿主的碱性蛋白酶重组菌的构建提供了新的思路。



 加载中...
加载中...
