


 13720223167
13720223167



众所周知,传统诱变育种包括诱变和筛选两部分,是获得优良突变体的有效途径之一.诱变部分包括出发菌株的选择、诱变剂种类和剂量的选择以及合理的使用方法.筛选部分包括初筛和复筛,选出产量高、性状优良的突变株,并且找出发挥此变株特性的最佳培养基及培养条件,使其在最适的环境条件下合成有效的产物[1,2].整个诱变育种过程复杂繁琐、效率低下.尤其体现在:
1.要以适宜致死率为指标对诱变剂种类和剂量进行选择以确定诱变条件;
2.无论是随机筛选还是平板菌落预筛首先都要将诱变后的菌悬液以一定特征(形态突变、抗性突变、生理生化突变等)分离纯化,并且大量挑选单菌落(一般200株左右)以寻找少数正突变;
3.性状更佳的突变体很可能因不具备某项筛选特征而被遗弃,即经诱变后的优良突变株有可能并非任何形态突变、抗性突变、营养缺陷型突变或温敏突变等而被舍弃.
4.对选出的少数正突变再连续反复数代诱变筛选;
5.对最终选定的正变株进一步进行发酵条件优化[1].
整个过程具有较大的盲目性,工作量大.笔者首次提出的多组复合诱变株混合发酵选育技术有效地克服以上不利方面,提高了整体诱变育种效率.
γ-多聚谷氨酸[γ-Poly(glutamicacid)简称γ-PGA]是一种由微生物合成的对人体和环境无害的生物相溶性天然高分子,具有增稠、乳化、凝胶、成膜、保湿和粘接等功能,有着极高的经济效益和社会效益.作为首次尝试,笔者以γ-PGA生产菌BacilluslicheniformisATCC9945A作为原始出发菌株,采用多组复合诱变株混合发酵选育技术获得相应高产突变株.
1 材料和方法
1.1 材料
1.1.1 出发菌种
地衣芽孢杆菌(Bacilluslicheniformis)ATCC9945A,由江南大学生物工程学院提供.
1.1.2 培养基
包括斜面活化培养基,牛肉膏蛋白胨培养基,种子培养基,基础发酵培养基,按照文[3]的方法配制.
1.2 方法
1.2.1 多组复合诱变株混合发酵选育技术的整体思路
传统诱变育种以分阶段方式进行:在诱变阶段完成后立即对诱变菌分离纯化,选出其中的正变株,在筛选阶段完成后再进行培养基和培养条件的调整.而笔者打破传统诱变育种的思维定式,认为获得高产的决定因素包括两方面:一是菌体本身诱变后具有高产特性,二是有适宜其高产特性发挥的培养条件,两者是有机结合的整体系统,因此诱变育种时应将两方面同时进行,一步到位.在此思路指导下设计了一种快速有效筛选突变株的方法:最初诱变时,选取三组有较大差异的诱变条件平行诱变,保证能在最大范围内诱变出所需的γ-PGA高产菌株.然后将三组诱变后的菌种全部混合,以这种包含所有正突变和负突变的混合菌出发接种发酵,通过"复杂系统调控技术"对发酵条件进行调控[4],以终产量为指标,选出有利于混合菌中的优良正变株产γ-PGA的发酵培养条件.经过几次调控,只有当混合菌中高产突变株的高产特性得以发挥并逐步积累,γ-PGA的产量才会逐步提高,最终得到适合高产突变株的最佳发酵条件.以最佳发酵条件为基础对混合菌进行分离纯化,由于高产菌株在此发酵条件下具有生长优势,否则对培养基调控γ-PGA产量不会有较大幅度提高,因此只需挑取较少的单菌落即可筛选到优良高产菌株,降低了筛选的盲目性,减少了繁琐的筛菌过程,同时找出了适合高产菌株的发酵培养条件,大大提高了传统诱变育种的工作效率(图1).

1.2.2 复杂系统定向调控技术原理简述
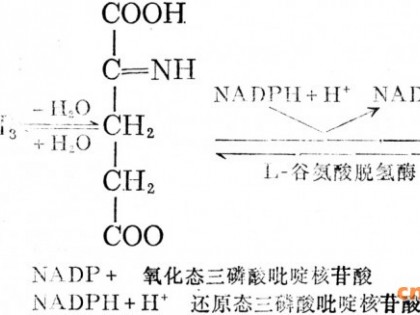
为了从整体上调控一个实际的复杂系统,首先必须把实际项目抽象为数值系统,应用数值化反馈技术,从而实现对复杂系统的调控.其原理如图2.Xi(i=1,2,...n)表示全部系统因素,它们分别代表物质和作用于物质的工艺因素.Fh(X1,X2,...Xn)表示全部系统因素之间的相互作用关系,数学上通常是用某种高阶非线性偏微分方程组表示.Y表示系统行为量值.A0表示系统行为的期望量值.当X1至Xn连续变化,则Y也随之变化.实际上对于复杂系统通常难以建立微分方程组,且即或建立但求其解很难,甚至不可能,因此着眼于实验、分析输入于输出间的相关性便成为复杂系统调控的基本思路,即分析Xi与Y之间的相关性及自变量X1,X2,X3,...Xn彼此之间的相关性.经由模糊逻辑判断及计算,形成N信道数值反馈控制,以修正自变量Xi,于是迫使系统行为Y迅速收敛于期望值A0.
1.2.3 诱变处理
采用紫外线(UV) 亚硝基胍(NTG)复合诱变[1] 将出发菌株以斜面活化试管活化,再以牛肉膏蛋白胨培养基扩大,使菌种位于对数生长期,离心洗涤菌体并以磷酸缓冲液调节菌落数为108个/mL左右.分3组以紫外线照射后再经亚硝基胍复合诱变,处理剂量及方式见表1.
经亚硝基胍处理后,将诱变后的各组菌体分别转入牛肉膏蛋白胨培养基中,在37℃下摇床6h以上,离心洗涤菌体两次以除去残留的亚硝基胍.将洗涤后的三组诱变菌混合一起,加以50%甘油水溶液分装保存于-70℃备用.
1.2.4 筛选平板的确立
对γ-PGA生产菌的发酵培养条件考察研究后[5,6],应用"复杂系统定向调控技术"设计调控筛选平板[4].将保存的混合突变株转入牛肉膏蛋白胨培养基活化,24h后,以不同的发酵条件开始发酵,见表2.测定各组发酵条件下的γ-PGA产量,再反馈调控各项影响发酵的因素,反复几次,最终以γ-PGA产量最高的一组培养基加入适量琼脂作为筛选平板,并将此组发酵条件定为最适发酵条件.
1.2.5 高产菌株的选育
将保存的混合突变株转入牛肉膏蛋白胨培养基活化24h后,转入种子培养基中,以最适发酵条件扩大.将发酵后的混合菌以梯度稀释涂已经筛选的平板.42h后挑取形态大、表面光滑、粘稠的单菌落进行纯培养[3].并以筛选出的菌株与出发菌平行发酵,选出γ-PGA产量最高的菌株.其中每株筛选菌皆采用我们优化调控后的最适发酵条件发酵.
1.2.6 遗传稳定性实验
将筛选出的高产菌株与出发菌株进行了平行继代遗传稳定性实验,共转接了10代,证明其是否是具有遗传稳定性的高产菌株.
1.2.7 γ-PGA产量的测定
将发酵液离心,取上清液加入4倍体积乙醇沉淀,搅拌过夜后,离心除去上清液,以蒸馏水将沉淀完全溶解并离心除去不溶性沉淀.测定此γ-PGA水溶液中谷氨酸单体含量,并以此γ-PGA水溶液加GR盐酸封管110℃水解24h,测定水解后的谷氨酸总量.两次测得的谷氨酸之差为γ-PGA含量[7,8].
1.2.8 谷氨酸含量的测定
采用纸层析法、比色定量法测定[9].
2 结果与分析
2.1 筛选平板的确定
X1~X14为物质条件(g/L),其中X1~X4为碳源,分别是柠檬酸、谷氨酸、甘油、麸皮汁.X5~X6为氮源NH4CL和NH4HCO3.X7~X14为金属离子,分别是K2HPO4、KNO3、Mg2+、Fe3+、Ca2+、Mn2+、Mo6+、Na+.X15~X21为工艺条件,分别是发酵温度(℃)、pH、装液量(mL)、接种量(%)、发酵时间(h)、转速(r/min)、种龄(h).Y为γ-PGA的产量(g/L).设计了3项抽样实验,即系统因素优化配置诊断性实验(表2).通过应用"复杂系统定向调控技术",将系统由原来的21因素简化为由16个因素组成的有效系统.通过进一步的调控实验,16因素的量值配比得到了最优化配置,致使系统行为(产量)达到最大的稳定.
通过对以上21因素组成系统诊断考察后,经由计算分析认为某些因素对系统而言属于多余组分,于是简化整个系统,形成新的16因素系统,对此16因素系统又进行进一步的诊断(表3).经分析考察系统行为,认为必须进一步实施调控,其试验设计与结果见表4.
通过以上实验结果可知,对于诱变后的混合菌而言,其中由正突变株产生的γ-PGA最高产量可达到3.847g/L,所以适合正突变株的最优配比培养条件如表4第一组所示.在这个量值最佳配比下,γ-PGA高产菌株能在有其他负突变株影响下获得最高产量.因此,我们有理由相信以此发酵条件作为筛选平板培养基的配方,对混合菌分离纯化,筛选出的正突变株在没有负变株作用的影响下,应取得极高的产量.
2.2 高产菌株的选育
从2.1确定的筛选平板上只分离纯化了40株形态大,表面光滑且粘稠的单菌落(Zγ-1~Zγ-40).将每个单菌落在2.1确立的最佳发酵条件下发酵后测定每株菌的γ-PGA产量(表5).而出发菌株BacilluslicheniformisATCC9945A的γ-PGA产量在使用了经调控后的最佳发酵条件培养也仅为4.582g/L.由此可见,由于培养条件向优势菌体倾斜,因而诱变后通过筛选平板筛选出的突变株中正突变占所有筛选菌的77.5%,平均产量都提高了3~4倍,大大降低了传统诱变育种时筛菌的盲目性,减少了工作量,提高了诱变效率.
2.3 遗传稳定性试验
我们将产量最高的5株突变株与出发菌ATCC9945A进行了平行继代遗传稳定性试验,每传代2次就测定一次γ-PGA产量,共转接10代,表6给出了产量最高的Zγ 29的遗传稳定性实验数据.
由表6可见,经诱变混合筛选出的Zγ 29具有稳定的遗传特性.同时其余4株具有较高产量的变株也都具有良好的遗传稳定性,数据不一一列出.
3 讨论
笔者针对产量仅为4.582g/L的γ-PGA生产菌BacilluslicheniformisATCC9945A施行了三组紫外线-亚硝基胍复合诱变,并将诱变后的三组菌混合发酵,并利用"复杂系统定向调控技术"确定最佳发酵条件,高效而准确地挑选到目标突变株Zγ 29.在最佳培养条件下对其发酵培养,γ-PGA产量高达18.99g/L,是出发菌株的4.14倍.本研究成果突出之处在于,有效地克服了传统诱变育种的5点不利方面,其表现是:
一.将三组不同诱变条件下的诱变菌全部混合,用于调控高产正突变的最适发酵培养基和发酵条件,完全省略了诱变阶段以致死率来对诱变剂种类和剂量的选择这一复杂的过程;
二.无需在诱变后立即进行大批量的正负突变株分离纯化筛选.
三.由于将所有诱变菌混合直接进行发酵条件调控,因而可以将诱变后的所有优良正变株包括,通过调控培养基和培养条件而发挥出其高产特性.而不会如传统诱变育种那样必须以形态突变或营养缺陷型突变株等特征来筛选遗漏了某些不具备这些特征的优良正变株.
四.无需经过几代连续诱变,即可达到目标要求.五.对高产菌的培养基和培养条件的优化早在筛选过程中完成,大大提高了诱变育种效率.值得一提的是,诱变后,我们对发酵条件进行调控的过程完全不同于传统诱变育种中对营养缺陷型或温度敏感突变型等变株筛选过程中的富集.后者仅是利用理化手段去除掉不具备该分离特征的菌株,使其相对量增加,而前者是以使γ-PGA产量提高为目的,对培养基和培养条件进行调控,在产量得以提高后我们可以假设这一提高与高产正突变菌的富集有一定关系,是正突变菌的绝对量增加,而后继的实验也证实了这一假设.因此相比之下,我们提出的技术思路在针对数量性状突变株的诱变育种方法上更具有目的性.
另外,我们通过对遗传稳定性实验结果分析表明进行遗传稳定性考察的5株产量最高的菌株都有良好的遗传稳定性,这在传统诱变育种中比较少见.这进一步证实了"多组复合诱变株混合发酵选育技术"思路的正确性.因为可能正是在我们对混合菌发酵条件的反复调控过程中,具有稳定遗传特性的优良正变株逐步发挥作用,才使γ-PGA产量得以提高.综上所述,本文提出的多组复合诱变株混合选育的微生物高效诱变育种新思路可能对其他微生物诱变育种有借鉴意义,具有重要的实用价值.
参考文献:[1] 施巧琴,吴松刚.工业微生物育种学(第二版)[M].北京:科学出版社,2003.
[2] 曹军卫,马辉文.微生物工程[M].北京:科学出版社,2003.
[3] 杨革,陈坚,曲音波,等.激光生物学报,2001,10(4):255-260.
[4] 邱凯,万昌秀,张兴宇,等.科技通报,2003,19(1):25-28.
[5] SungHoYoon,JinHwanDo,etal.BiotechnologyLetters,2000,22:585-588.




 加载中...
加载中...
