


 13720223167
13720223167



摘要 本文综述了利用微生物生产L-色氨酸的各种方法和L-色氨酸的生物合成途径及其代谢调控机制,并介绍了利用重组DNA技术选育L-色氨酸高产菌的研究现状。
L-色氨酸是含有吲哚基的中性芳香族氨基酸,为白色或略带黄色叶片状结晶或粉末,水中溶解度1.l4g(25℃),溶于稀酸或稀碱,在碱液中较稳定,强酸中分解。微溶于乙醇,不溶于氯仿、乙醚。它是人体和动物生命活动中必需的氨基酸之一,对人和动物的生长发育、新陈代谢起着重要的作用,被称为第二必需氨基酸,广泛应用于医药、食品和饲料等方面。在生物体内,从-色氨酸出发可合成5-羟基色胺等激素以及色素、生物碱、辅酶、植物激素等生理活性物质,可预防和治疗糙皮病,同时具有消除精神紧张、改善睡眠效果等功效。色氨酸代谢失凋会引起糖尿病和神经错乱,因此在医学上被用作氨基酸注射液和复合氨基酸制剂。另外,由于色氨酸是一些植物蛋白中比较缺乏的氨基酸,用它强化食品和做饲料添加剂对提高植物蛋白质的利用率具有重要的作用,它是继蛋氨酸和赖氨酸之后的第三大饲料添加氨基酸。
1.色氨酸的生产方法
色氨酸的生产最早主要依*化学合成法和蛋白质水解法,但是随着对微生物法生产色氨酸研究的不断深入,这种方法已经走向实用并且处于主导地位。微生物法大体上可以分为直接发酵法、微生物转化法和酶法。近年来还出现了将直接发酵法与化学合成法相结合、直接发酵法与转化法相结合生产色氨酸的研究。另外,重组DNA技术在微生物育种和酶工业上的应用极大地推动了直接发酵法和酶法生产色氨酸的工业化进程。
1.1微生物转化法
亦称前体发酵法。这种方法使用葡萄糖作为碳源,同时添加合成色氨酸所需的前体物如邻氨基苯甲酸、吲哚等,利用微生物的色氨酸合成酶系来合成色氨酸。这种方法同直接发酵法一样,需要解除生物合成途径中大部分酶所受到的反馈调节,以使色氨酸能够高浓度蓄积。另外,所添加的前体物大都是抑制微生物生长的,因此添加量不可过高,一般采取分批少量添加的方法。同时可以筛选前体物的抗性突变株来提高前体物的添加量。例如采用枯草杆菌的5-FT抗性突变株SD-9在15L含6%葡萄糖的培养基中培养,在培养过程中每次添加少量6%的邻氨基苯甲酸溶液共2L,经120h后可生产色氨酸9.6g/L。微生物转化法的不足在于当转化液中前体物浓度较高时,转化率有所下降。另外,前体物的价格比较昂贵,不利于降低成本。
1.2酶法
酶法是利用微生物中色氨酸生物合成酶系的催化功能生产色氨酸。这些酶包括色氨酸酶、色氨酸合成酶、丝氨酸消旋酶等。根据提供这些酶的微生物种类数,可以分为双酶菌法和单酶菌法两种类型。该法既可以直接加入细胞壁溶解酶使细胞破壁后再使用,也可以将所需的酶固定化后再使用,一般由酶源菌体的培养、菌体的分离洗涤、固定化和反应几个阶段组成。例如利用大肠杆菌的色氨酸合成酶和恶臭假单孢菌的丝氨酸消旋酶,以吲哚和DL-丝氨酸为底物,在200L反应罐中反应24h,色氨酸产量可达到28.5g/L。利用黄杆菌AF39l2的酶系,以DL-5-吲哚甲基乙丙酚脲为底物,同时在反应液中加入适量的苯肼或野芝麻花碱以抑制色氨酸氧化酶的活性而阻止色氨酸的降解,L-色氨酸的产量可达8.8g/L。酶法能够利用化工合成的前体物为原料,既充分发挥了有机合成技术的优势,又具有产物浓度高、收率高、纯度高、副产物少、精制操作容易的优点,是一种成本较低的生产色氨酸的工业化生产方法。
1.3直接发酵法
该法是以葡萄糖、甘蔗糖蜜等廉价原料为碳源,利用优良的色氨酸生产菌种来生产色氨酸。对这种方法的研究进行的比较早,但在相当长的一段时间内达不到工业化生产的要求。主要原因是从葡萄糖到色氨酸的生物合成途径比较漫长,其代谢流也比较弱,而且色氨酸的合成需要多种前体物(如PRPP等),若想进一步提高色氨酸的积累量就必需设法增强合成这些前体物的代谢流。另一方面,色氨酸生物合成途径中的调控机制比较复杂,除了反馈调节这一粗调系统之外,还存在着细调系统-弱化子系统。随着重组DNA技术在微生物育种中的应用,为优良的色氨酸生产菌株的筛选和产酸水平的提高提供了可*的技术保障,使微生物直接发酵法生产色氨酸成为一种廉价的工业化生产方法。例如从大肠杆菌EMS4-C25中分离得到抗反馈调节的色氨酸操纵子,并将其克隆到质粒pUCI9和pHSG576中分别得到重组质粒pTC7O1和pTC576。将pTC576转化到大肠杆菌中,该菌经过17h的分批发酵积累色氨酸l6g/L。Ikeda等将带有DAHP合成酶(DS)和色氨酸合成酶(F)的质粒引入产色氨酸的谷氨酸棒杆菌KYl0-894中,使色氨酸的产量提高了54%,达到了66g/L。
l.4 其他生产方法的研究
这些方法主要是将上述几种方法有机地结合起来进行色氨酸的生产。主要有直接发酵法与化学合成法、酶法相结合,其特点是利用发酵法廉价提供一种前体物,再结合其它方法的优势进行生产。例如,利用黄色短杆菌P390直接发酵积累L-谷氨酸-y-半醛(GSA)达13.2g/L,然后将发酵液适当稀释后加入含苯肼的1mol/L H2SO4中加热回流1h之后,48%的CSA可转化为 L-色氨酸。利用落叶松草AHU9382以5%葡萄糖为碳源发酵3d可积累丙酮酸22-26.5g/L,然后加入吲哚、氯化铵和具有高色氨酸酶活性的产气肠杆菌培养,12h之后可积累L-色氨酸l5g/L。
2.色氨酸发酵的代谢控制育种
近年来,色氨酸作为必需氨基酸在输液、健康食品、饲料添加剂上的应用日益广泛,随着蛋氨酸、赖氨酸的大量饲料化应用,价格一直较高的色氨酸更希望有廉价制造的工业化生产方法,以便推进色氨酸的应用饲料化的实施。在我国对直接发酵法生产色氨酸虽作了一些研究工作,但发酵产酸水平较低,还没有得到可应用于工业化生产的稳定的高产菌株,因此,研究色氨酸的生物合成途径及代谢调节机制,用代谢控制育种和基因工程的方法构建色氨酸高产菌株就十分必要。
2.1出发菌株的选择
色氨酸发酵的生产菌种有谷氨酸棒杆菌(C·glutamicum)、黄色短杆菌(B·flavum)、枯草芽孢杆菌(B·subtilis)、大肠杆菌(E·coli)、产朊假丝酵母(Candida utilis)等。
2.2色氨酸的生物合成途径和代谢调节机制
迄今被研究的所有菌株的色氨酸生物合成途径都是一样的,但是其调控方式及trp基因的表达等却有很大的不同。这些不同点主要有:
(1)DS酶所受的调节方式不同。在大肠杆菌、粗糙脉孢菌等微生物中有3种DS酶,这些同功酶分别受3种芳香族氨基酸的反馈调节。在谷氨酸棒杆菌和黄色短杆菌中只有一种DS酶,它受L-Phe和L-Tyr的协同反馈抑制。枯草芽孢杆菌中的DS酶受顺序反馈抑制。
(2)和色氨酸生物合成有关的基因的顺序和结构不同。在肠杆菌中对色氨酸的合成进行调控的基因组成一个单一的操纵子,而且它们排列的顺序也都是一样。在乳糖发酵短杆菌中也是单一的操纵子结构,但在非肠杆菌和真核生物中这种结构却并不多见。
(3)对结构基因表达的调控方式不同。在大肠杆菌等肠杆菌中,依*阴遏系统和转录的弱化机制来对和色氨酸生物合成有关的结构基因的表达进行调控,在枯草芽孢杆菌中需要RegA和mtrB基因来参与对转录弱化的调控和RNA结合蛋白的调节。在铜绿假单孢菌中,trpI基因编码的转录激活因子在吲哚甘油磷酸存在时诱导rtpB、trpA基因的表达。在有些微生物中,只有转录的弱化机制对基因的表达进行调控。
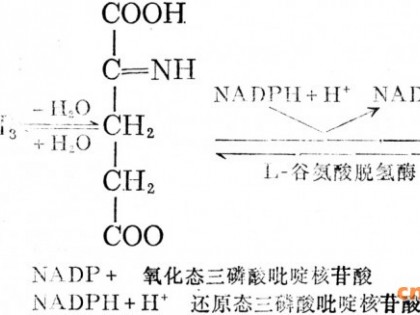
色氨酸的生物合成途径和代谢调节机制(以黄色短杆菌为例),邻氨基苯甲酸合成酶(AS)被色氨酸反馈抑制1.5umol/L的Trp使其活性丧失50%,抑制常数Ki为0.44umol/L,10umol/L的Try 几乎完全抑制其活性,这种抑制作用大大强于大肠杆菌和枯草芽胞杆菌中的AS酶所受的抑制作用。对于AS酶的底物分支酸来说,抑制是竞争性的,而对于另一种底物谷氨酰胺,抑制是反竞争性的。邻氨基苯甲酸磷酸核糖移换酶(PRT)不与AS形成复合体,主与大肠杆菌有所不同。它也受Trp的反馈抑制,其抑制常数Ki为0.26mmol/L。对于该酶的两个底物邻氨基苯甲酸核糖焦磷酸来说,抑制都是非竞争性的。TS酶乳受Trp的反馈抑制,其抑制常数为2.0mmol/L,约比PRT的Ki高一个数量级,对于其底物丝氨酸来说,抑制是竞争性的。此外色氨酸专一合成途径中的所有酶均受到Try的反馈阻遏,但它们的活性系数各不相同,显示出不同程度的阻遏效应。
对于色氨酸操纵子的结构及其阻遏系统和弱化机制已经研究得比较清楚,本文对此不再赘述。值得注意的是,在色氨酸操纵子内部还存在一个二级启动子,该启动子位于trpC基因的上游、TrpD基在的末端。为什么trp操纵子需要两个转录起始位置,对这个问题还没有明确的答案,但这很可能是对Trp生物合成进行调节的经济性的需要。由于Trp对TrpE、TrpD所编码的邻氨基苯甲酸合成酶(AS)和邻氨基苯甲酸核糖移换酶(PRT)的抑制作用远大于对TrpC、TrpB、TrpA所编码的3个酶的抑制作用,因此没有必要使这5个酶的合成具有同步性。色氨酸操纵子中的两个启动子具有不同的强度,启动作用受不同的因子调控,可使基因表达更加有效、更加协调,而且在不同的生活环境中,不同的启动子精密地调节基因的表达量,对维持细胞的生存起着非常重要的作用。
3.色氨酸生产菌的育种思路
如上所述,芳香族氨基酸的生物合成存在着特定的代谢调节机制,因此不可能从自然界中找到大量积累色氨酸的菌株。但是我们可以以黄色短杆菌、谷氨酸棒杆菌等为出发菌株,设法得到从遗传角度解除芳香族氨基酸生物合成证常代谢调节机制的突变菌株,用微生物直接发酵法,生产色氨酸。这些方法包括:解除菌体自身反馈调节,切断支路代谢,增加前体物的合成等,下面分别讨论。
3.l切断支路代谢
切断由分支酸到预苯酸、Vk、CoQ的代谢支路,可节约碳源,使中间产物分支酸更多地转向合成色氨酸,而且还可以解除苯丙氨酸、酪氨酸对合成途径中DS的反馈调节。从而有利于色氨酸积累。例如,有人以谷氨酸棒杆菌为出发菌株,经诱变得到的KY9456(Phe+Tyf)可积累色氨酸0.15g/L。这个产量是很低的,由此也可看出只选育缺陷型是不够的,还必须解除由色氨酸引起的反馈调节。
3.2解除菌体自身反馈调节
根据图1所示的调节机制,对于黄色短杆菌来说,如果解除色氨酸特异途径的调节机制,即使有共同途径上反馈调节的存在,也能大量积累色氨酸。因此可选育色氨酸结构类似物的抗性突变株,解除其自身的反馈调节来达到积累色氨酸的目的。对于色氨酸来说,它的结构类似物有:5-甲基色氨酸(5-MT)、5-氟色氨酸(5-FT)、色氨酸氧肟酸盐(TrpHx)、6-氟色氨酸(6-FT)、6-甲基色氨酸(6-MT)、4-甲基色氨酸(4-MT)等。它们都可以由于结构与色氨酸类似而被AS酶误认,与AS酶的调节部位结合。它们的抗性突变株就可以解除色氨酸对AS酶及该特异途径中其它酶的反馈调节,从而使色氨酸得以积累。此外,选育分支酸的结构类似物重氮乙酰丝氨酸(AZ)抗性突变株也能有效地提高AS酶的活性,从而提高色氨酸的积累量。
3.3增加前体物的合成
为了积累更多的色氨酸,必须更多地增加前体物,其中包括减少PEP和EP的支路代谢,解除苯丙氨酸和酪氨酸对DS的反馈调节,增加分支酸的浓度等方法。为了积累更多的PEP,防止丙酮酸生成更多的草酚乙酸,可以选育磷酸烯醇式丙酮酸羟化酶和丙酮酸激酶活力低的菌株或者选育硫辛酸和硫胺素(分别为丙酮酸脱羧酶系和丙酮酸羧化酶系的辅酶)双重缺陷型突变株。
解除苯丙氨酸和酪氨酸对DS的反馈调节,除了选育它们的营养缺陷型外,还需选育苯丙氨酸和酪氨酸结构类似物的抗性突变株,从而使DAHP得以大量合成,进而生成分支酸,并在此处优先合成色氨酸。它们的结构类似物有对氟苯丙氨酸(PFP),苯丙氨酸氧肟酸盐(PheHx),p-2-噬嗯基丙氨酸,对氨基苯丙氨酸(PAP),3-氨基酪氨酸(3-AT),酪氨酸氧肟酸盐(TyrHx),D-酪氨酸等。此外,选育磺胺胍抗性突变株也可以有效地提高分支酸的浓度,在具有前述一些标记的基础上增加这些抗性标记,可使色氨酸的产量进一步提高。
3.4切断进一步代谢
选育色氨酸酶(TN)缺失突变株,色氨酸脱羧酶缺失突变株,色氨醚tRNA合成酶缺失突变株以及不分解利用色氨酸的突变株,可以减少色氨酸的消耗,从而有利于色氨酸的积累。
3.5利用基因工程技术构建色氨酸工程菌
以上所述的诱变育种方法,主要在于解除合成能力强大菌株的代谢调节机制,令其积累目的产物。而基因工程技术,则主要在于通过基因扩增,增强生物合成途径中的限速酶或转换酶,这是提高色氨酸产量的有效措施,也是今后育种的主要方向。其方法主要有:(1)对编码Trp生物合成途径中的限速酶的基因如DS酶基因、TrpE、TrpD、TrpC、TrpB、TrpA等进行扩增。Hershfield等人采用ColEl质粒,将来自80dt的色氨酸操纵子克隆到大肠杆菌中,随着每细胞拷贝数增加20-30,色氨酸合成酶的活性增加150倍。这种酶活性的增加,必然反映在终产物色氨酸的积累量上。日本的Katsumata Ryoichhi等人从谷氨酸棒杆菌K86中获得质粒pCDTrpl57,该质粒带有DS酶基因和TS酶基因。将该质粒转化到谷氨酸棒杆菌KY9182(Phe-),得到KY9T82/pCDtp157,该菌可产色氨酸1.1g/L。将与Trp合成有关的基因克隆到质粒pDB9901上,然后导入Trp甲生产菌BRS-13中,在500ml培养基中30℃通风培养,可产色氨酸35.2g/L(亲株产20.1g/L)。(2)对某些酶的基因进行体外诱变造成该基因所编码的酶不可逆失活,再将突变基因导入色氨酸生产菌中。这些基因包括色氨酸操纵子上的结构基因、TrpR、TrpA、TrpA)pheA等,它们发生突变后可有效解除Trp的反馈调节或切断分支代谢从而有利于色氨酸的积累。(3)对于其它基因的扩增或改造。这种方法主要针对已有一定产量基础的工程菌株,在这些菌株中,合成Trp的限速步骤不再是AS酶、DS酶等引起的(编码它们的基因已被扩增或发生突变),而多是由于前体物供给量不足、葡萄糖代谢流的不平衡等原因造成的。这些基因包括pgi、serA、PRPP合成酶基因等。日本的Yajima Yoshihiro等人从色氨酸生产菌(枯草杆菌)中分离出含PRPP合成酶的基因片段,将其克隆到质粒pSEY146112上,在促进剂p201的调节下,将其导入具有5-FT的枯草杆菌中,转化后PRPP合成酶的活力比原菌高提3.9倍。在含邻氨基苯甲酸800ppm时发酵,产色氨酸量比原菌提高1-l.25倍。利用基因工程技术使大肠杆菌K-12分别带上有缺失的pgi基因(编码磷酸葡萄糖异构酶)和正常的pgi基因,对照培养后发现pgi菌株从葡萄糖合成色氨酸的能力比pgi+高出近两倍。这说明重新分配葡萄糖在EMP和HMP途径中的代谢流,使RMP途径得到加强后,将加速NADPH和一些重要的戊糖衍生物的合成,提高色氨酸生物合成途径中的能量和前体物供给水平而使色氨酸积累量上升。Ikada等人将丝氨酸代谢途径中的磷酸甘油酸脱氢酶(PGD)的基因克隆到质粒pWK99中,构建出重组质粒pwK9901,将后者转入色氨酸生产菌KY10-894中(产酸钙以U,解决了该菌株在发酵过程中由于丝氨酸供给量较少而积累副产物吲哚,导致发酵后期耗糖减少、糖代谢受阻的问题。
3.6 其他标记
为了更多地生成色氨酸,可采用增加细胞膜渗透性的方法,使细胞内的色氨酸向培养基中渗透,降低细胞内的色氨酸含量,促使反应向生成色氨酸的方向进行。这可借助于谷氨酸生产菌选育时常用的方法,如选育Vp类衍生物抗性突变株,溶菌酶敏感突变株,甘油缺陷突变株,油酸缺陷突变株等。




 加载中...
加载中...